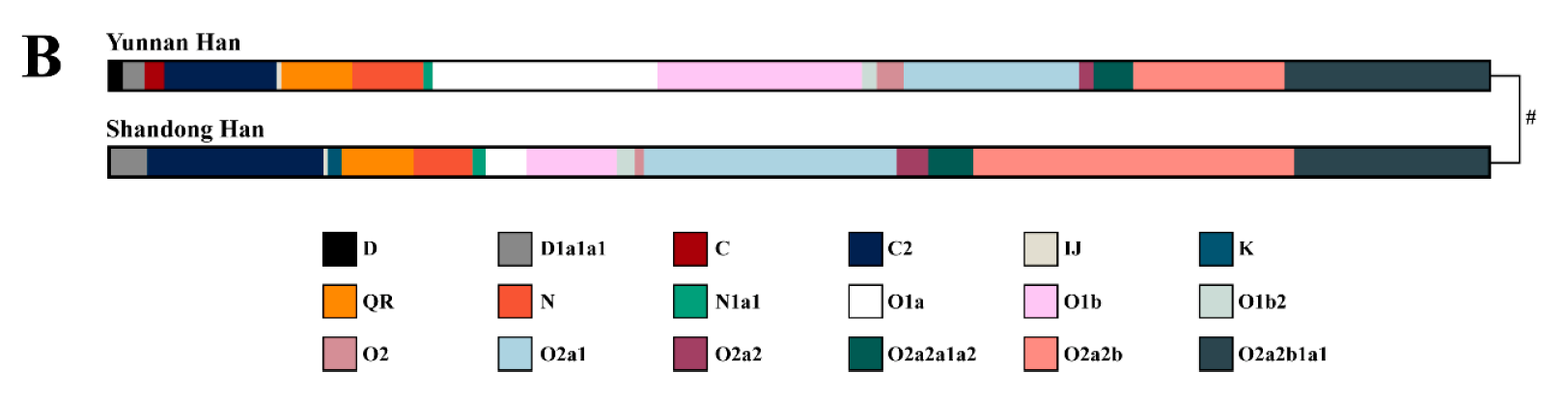
日本人によく見られるY染色体DNAのO系統、D系統、C系統については、すでに以下の記事を書きました。
- 日本人のY染色体ハプログループOの研究、人と稲作と言語の広がりは必ずしも一致しない
- アイヌ人と沖縄人のDNAを比べると・・・(Y染色体ハプログループDの研究)
- アフリカから始まる人類の歴史(Y染色体ハプログループCの研究)
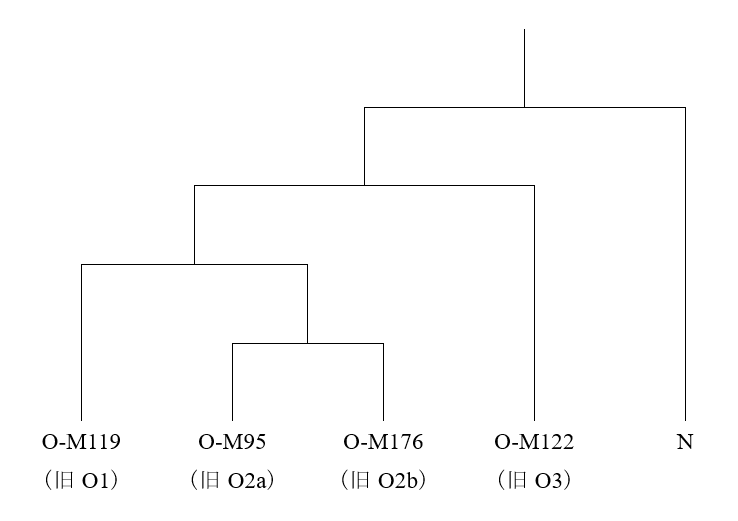
日本人のY染色体DNAは、基本的に以下の通りです。
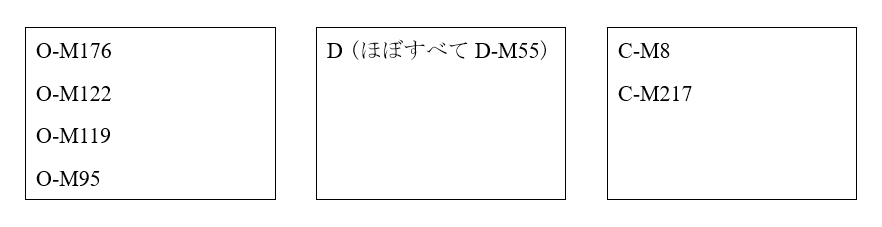
※まれにN、Q、Rが見られることがあります。
このうちのDとC-M8は、日本の近隣にほとんど見られず、縄文時代の日本列島に存在していたと考えられる系統です。したがって、弥生時代のはじめに日本列島に入ってきた人々のY染色体DNAは、O-M176、O-M122、O-M119、O-M95、C-M217です。
O-M176、O-M122、O-M119、O-M95は、直前の三つの記事で論じたように、農耕(イネ、アワ、キビの栽培)と密接な関係のある系統です。C-M217は、モンゴル系言語の話者とツングース系言語の話者に多く見られる系統で、もともと農耕との関係が薄い系統です。
モンゴル系言語の話者とツングース系言語の話者に多く見られるC-M217は、現代の日本人に数パーセントしか見られません(Hammer 2006、Nonaka 2007)。弥生時代のはじめに日本列島に入ってきた人々は、イネ・アワ・キビの栽培を日本に導入した人々であり、それらの人々に、農耕と密接な関係のあるO(O-M176、O-M122、O-M119、O-M95)が多く、農耕との関係が薄いC-M217は少なかったというのは、うなずける話です。
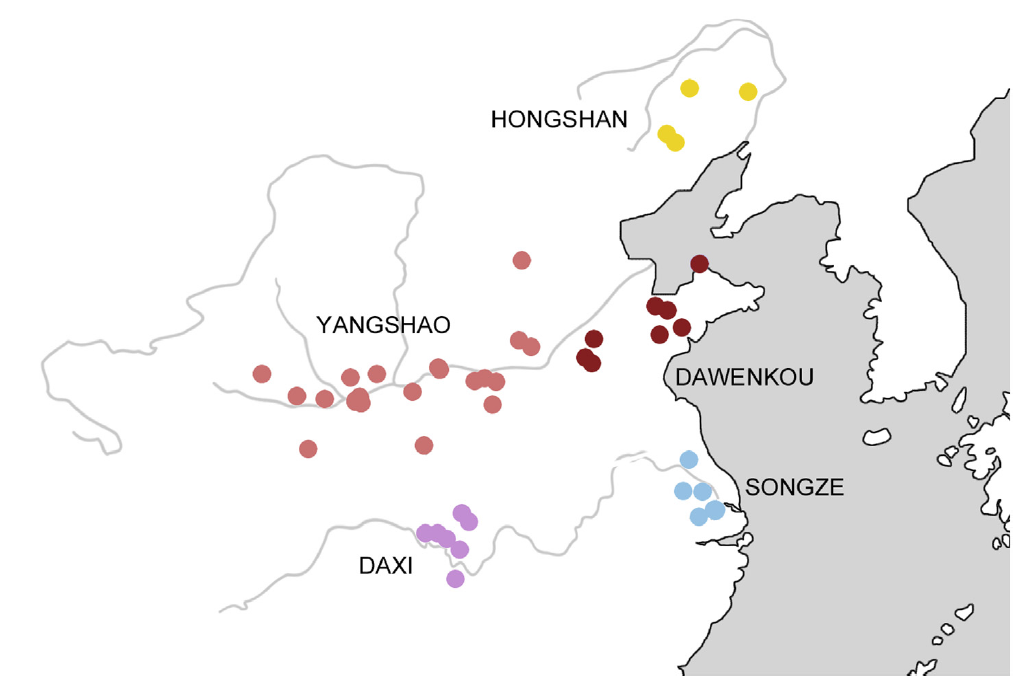
ただ、日本語の起源という観点からすると、これはなかなか複雑怪奇な話です。以下は、本ブログで何回も引用しているRobbeets 2021の図です。
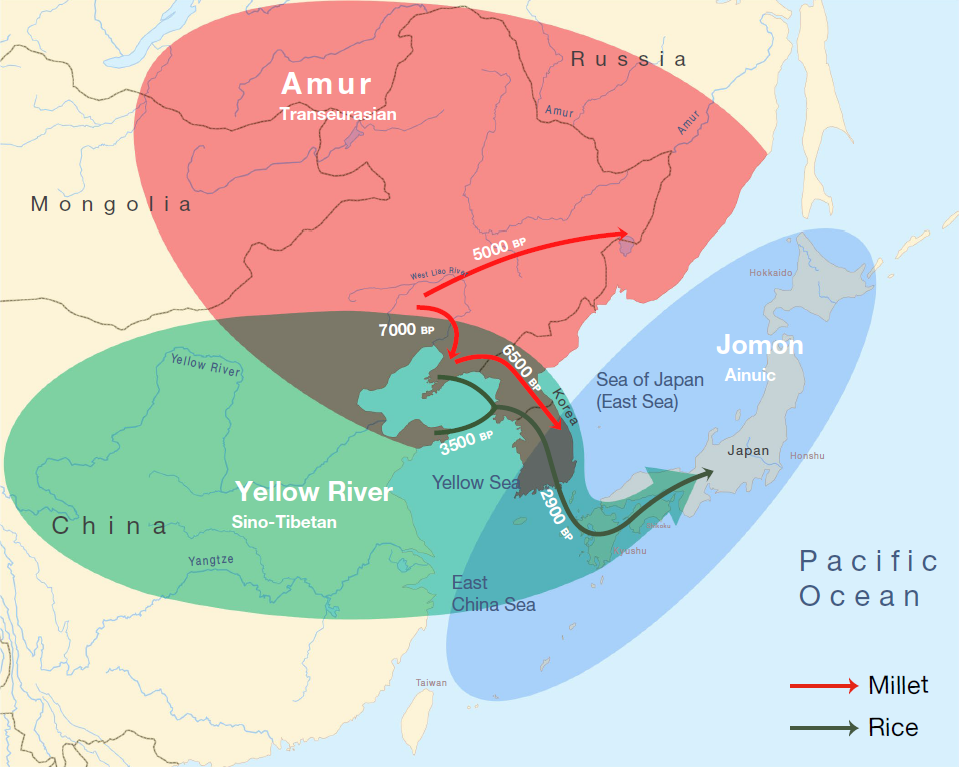
Robbeets氏らは、日本語は朝鮮語、ツングース諸語、モンゴル諸語、テュルク諸語と近い系統関係があるという旧態依然とした「アルタイ語族仮説」(トランスユーラシア語族と改称されていますが、実体は変わっていません)を主張しており、この点で、筆者の考えは全然違います。しかし、日本語が遼河流域から来ていると考える点では、Robbeets氏らも筆者も同じです。
Robbeets氏らの研究では、黄河流域~アムール地方の古代人のゲノム(DNA全体)を調べ、黄河流域の古代人を一方の極、アムール地方の古代人を他方の極として、遼河流域の古代人が中間的な存在であることを明らかにしています(Robbeets 2021)。これは、別の研究でも同様の結果が出ており、間違いありません(Ning 2020)。
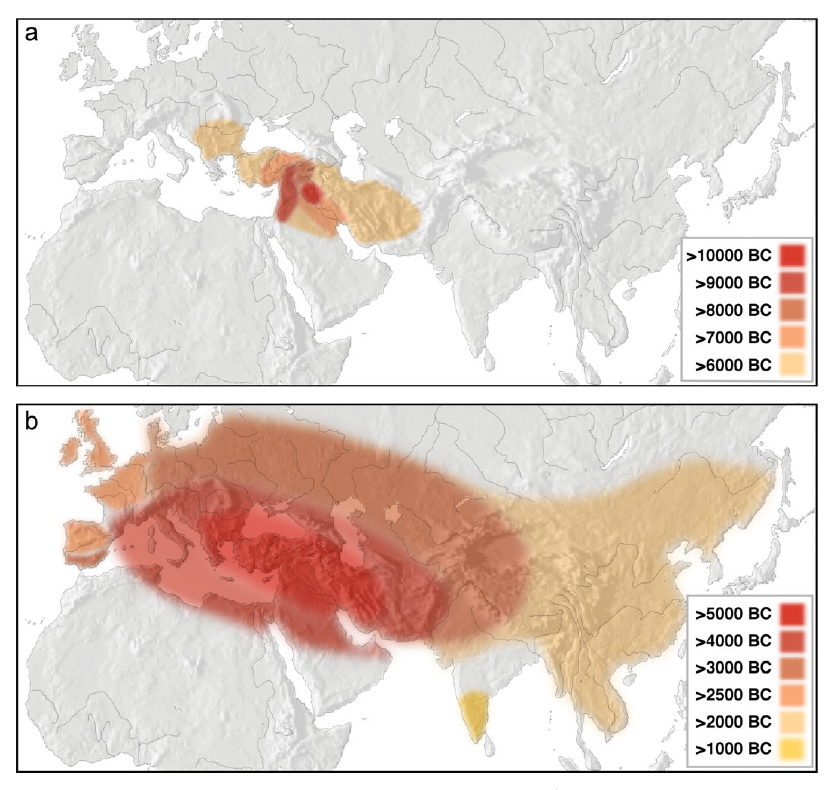
Robbeets 2021は言語学と生物学(遺伝学)と考古学を総合した研究ですが、Ning 2020は生物学(遺伝学)に焦点を当てた研究です。以下の図は、Ning 2020からの引用です。
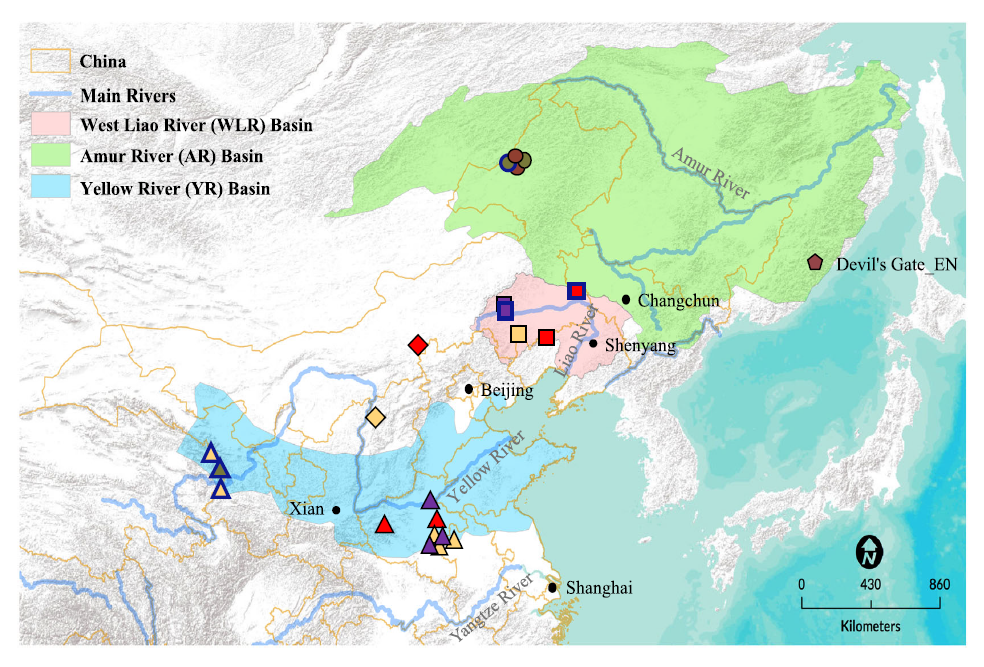
Ning氏らの研究は、「黄河流域」、「遼河流域」、「アムール地方」と、三つの地域に区分して古代人のDNAを調べているのが大きな特徴です。Robbeets 2021は、ゲノム(DNA全体)を調べた研究であり、それはそれで価値があるのですが、Y染色体DNAには焦点を当てていません(Y染色体DNAはゲノム(DNA全体)の一部ですが、Y染色体DNAは男性しか持っておらず、ひたすら父から息子へ受け継がれていくという特殊な性質を持っています。そのため、Y染色体DNAが示す傾向はゲノム(DNA全体)が示す傾向と大きく異なることがあり、Y染色体DNAは別個に調べる必要があります)。古代の遼河流域では、黄河方面のO-M122ともアムール方面のC-M217とも違うN系統が支配的だったのです(Cui 2013)。そして、遼河流域と黄河流域の間の地域では、NとO-M122が隣接していたのです(Zhang 2017)。
遼河文明の変遷を思い出してください(波乱の時代の幕開け、崩れゆく山東龍山文化、そしてそこに現れた異質な岳石文化を参照)。興隆窪文化(こうりゅうわぶんか)→趙宝溝文化(ちょうほうこうぶんか)/富河文化(ふがぶんか)→紅山文化(こうさんぶんか)と発展しましたが、5000年前頃から気候が悪化し始め、衰退します。4200~4000年前頃に、どん底まで落ちます。その後、持ち直して、4000~3500年前頃には夏家店下層文化(かかてんかそうぶんか)が栄えました。
紅山文化が栄え、衰退期があり、夏家店下層文化が栄えたわけですが、この間に遼河流域のDNAは大きく変化しました。
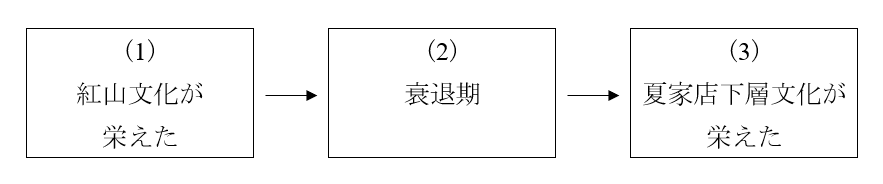
Robbeets 2021のゲノム分析でも、Ning 2020のゲノム分析でも、(1)の時代のDNAがそのまま(3)の時代のDNAになったのではなく、黄河方面のDNAがなだれ込んできたことが示されています。単純に、(1)の時代の人々が、(2)の時代を耐え、(3)の時代の人々になったわけではないということです。
Cui 2013のY染色体DNA分析でも、同様の傾向が示されています。人数は少ないですが、(1)、(2)、(3)の各時代を調べています。(1)の時代にはNが支配的で、(2)の時代にもNが支配的だったが、どん底を過ぎた後の(3)の時代ではO-M122がNに迫っているのが窺えます。
上の(1)→(2)→(3)は、遼河流域のみの観察です。(2)から(3)に向かう時期に、黄河方面のDNAの顕著な流入が認められます。遼河流域にいる人間集団をずっと観察していても、そうなのです。(2)の苦しい時代に、農耕を続けようとして、南に下っていった人間集団もいたはずです。その人間集団はどうなったのでしょうか。
日本語が遼河流域から山東省に南下した言語であると見られることは、以下の記事でお話ししました。
日本語(正確には日本語の前身言語)を話していた人間集団とは、まさに上に述べた遼河流域から南下していった人間集団であり、この人間集団がどうなったのかという問題は、日本語の歴史を考えるうえで最も重要な問題なのです。
(2)から(3)に向かう時期に、遼河流域にいた人間集団は、外(黄河方面)からの強い影響にさらされました。遼河流域から南下していった人間集団は、外からの強い影響にもっとさらされたにちがいありません。そんな状況の中で、遼河流域から南下していった言語はどうやって生き残ったのかと考えたくなるところです。次回の記事では、この問題について具体的に考えます。
これまでに蓄積されたデータから、かつての遼河流域でY染色体DNAのN系統が支配的であったことは確実です。その一方で、山東省から朝鮮半島、さらに日本列島に渡った人間集団で、Nではなく、O-M122とO-M176が支配的であったことも確実です(朝鮮半島と日本列島にO-M119とO-M95がわずかしか見られないことはすでに述べました(Nonaka 2007、Kim 2010))。日本語(正確には日本語の前身言語)を話す人間集団のY染色体DNAがなぜ(あるいはどのように)NからO-M122とO-M176に入れ替わったのかという問題に直面することになります。稲作の広がりとDNAの広がりが一致しないケースをすでに見ましたが、今度は言語の広がりとDNAの広がりが一致しないケースを見ることになります。
参考文献
Cui Y. et al. 2013. Y Chromosome analysis of prehistoric human populations in the West Liao River Valley, Northeast China. BMC Evolutionary Biology 13(1): 216.
Hammer M. F. et al. 2006. Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes. Journal of Human Genetics 51(1): 47-58.
Kim H. et al. 2010. Y chromosome homogeneity in the Korean population. International Journal of Legal Medicine 124(6): 653-657.
Ning C. et al. 2020. Ancient genomes from northern China suggest links between subsistence changes and human migration. Nature Communications 11: 2700.
Nonaka I. et al. 2007. Y-chromosomal binary haplogroups in the Japanese population and their relationship to 16 Y-STR polymorphisms. Annals of Human Genetics 71(4): 480-495.
Robbeets M. et al. 2021. Triangulation supports agricultural spread of the Transeurasian languages. Nature 599(7886): 616-621.
Zhang Y. et al. 2017. Genetic diversity of two Neolithic populations provides evidence of farming expansions in North China. Journal of Human Genetics 62(2): 199-204.