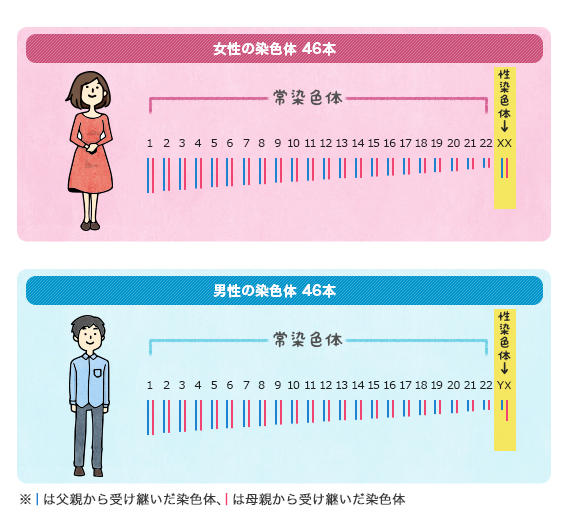

デニソワ人がロシアのアルタイ地方のデニソワ洞窟で発見されたのは、2008年のことです。ちょうど古代人の骨や歯からDNAを調べる技術が確立し始めたところで、デニソワ人の発見は実にタイムリーな出来事でした。それ以来、デニソワ人の注目度はぐんぐん上がっています。
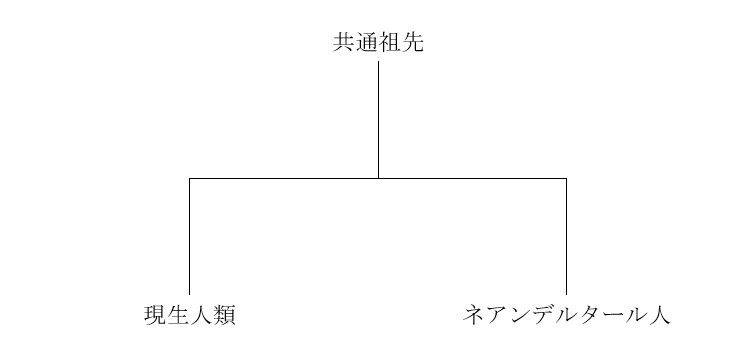
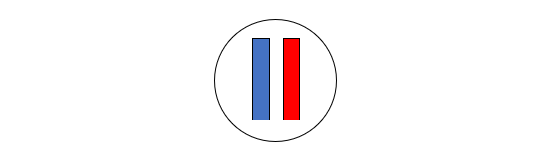
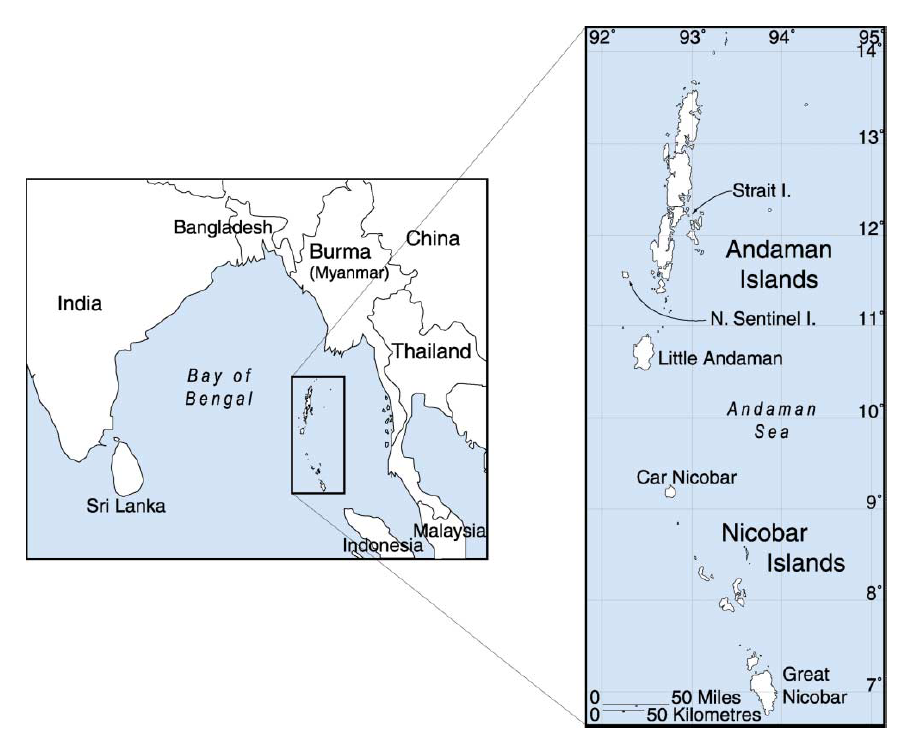
前回の記事で述べたように、まずデニソワ人のミトコンドリアDNAが調べられ、デニソワ人が現生人類とネアンデルタール人に近い種であることが明らかになりました(Krause 2010)。ミトコンドリアDNAを調べた段階では、現生人類とネアンデルタール人とデニソワ人は以下のような関係にあると見られました。
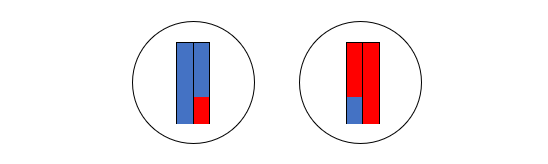
図1
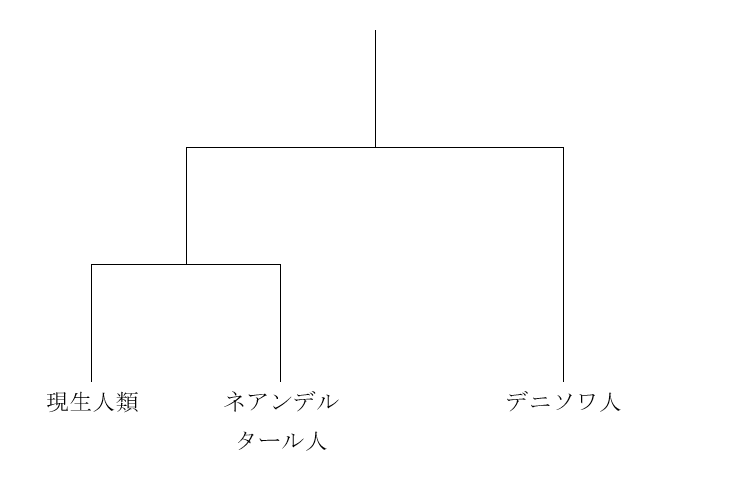
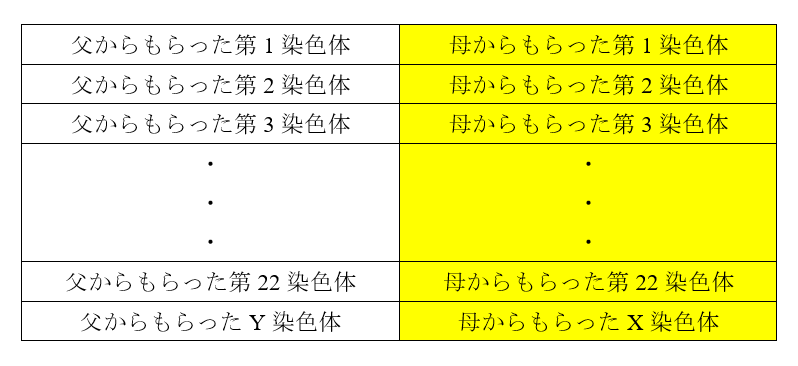
図1は、現生人類とネアンデルタール人が互いに近い関係にあり、デニソワ人が両者から離れた関係にあることを示しています。これはあくまで、ミトコンドリアDNAに基づく構図です。DNAと減数分裂の仕組み、ミトコンドリアDNAとY染色体DNAからは窺い知れない歴史の記事で説明したように、私たちのDNAのほとんどは、ミトコンドリアではなく、核にあります。ミトコンドリアDNAだけでなく、肝心の核DNA(=常染色体(第1染色体~第22染色体)DNA+X染色体DNA+Y染色体DNA)を調べなければなりません。上記のKrause氏らの研究グループは、今度はデニソワ人の核DNAを調べました(Reich 2010)。すると、ミトコンドリアDNAの時とは違う結果になりました。
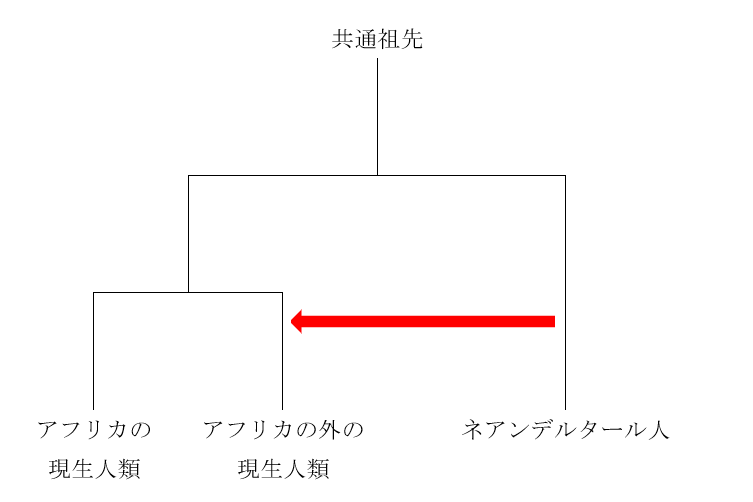
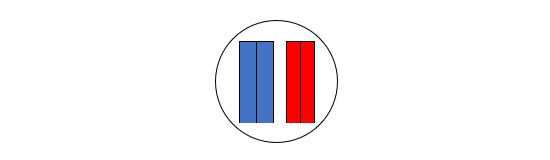
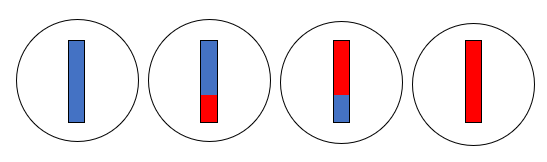
図2
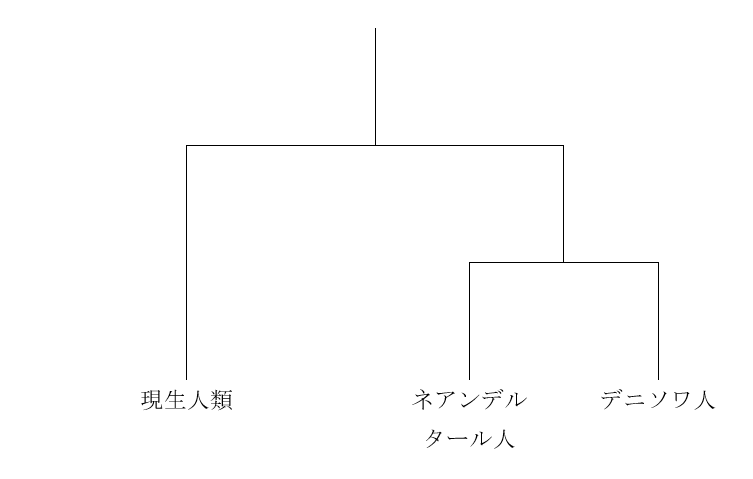
図2は、ネアンデルタール人とデニソワ人が互いに近い関係にあり、現生人類が両者から離れた関係にあることを示しています。要するに、ミトコンドリアDNAを調べると、現生人類とネアンデルタール人が近いという結果が出て、核DNAを調べると、ネアンデルタール人とデニソワ人が近いという結果が出たわけです。
私たちのDNAのほとんどは核DNAであり、核DNAのデータのほうが歴史の大まかな展開をよく表していると考えられます。実際の歴史は図2に近そうです。しかし、ミトコンドリアDNAを調べた時の図1の結果も無視できません。実際の歴史は図2に近いが、図2とは少し異なるのかなと考えたくなります。実は、核DNAのデータとミトコンドリアDNAのデータが食い違うケースはほかにもあるのです。以下の図を見てください。
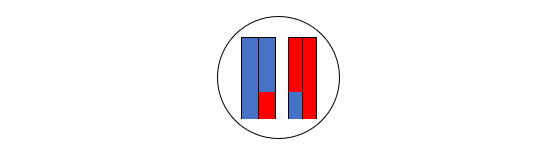
図3
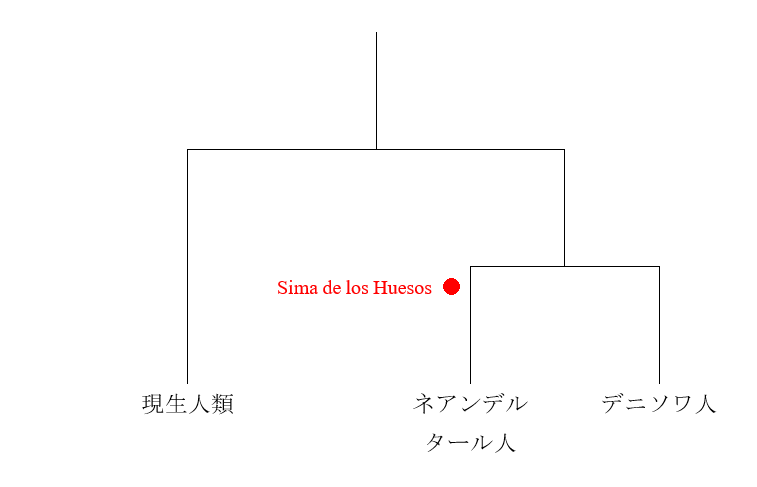
Sima de los Huesosと書き入れました。Sima de los Huesosというのは、スペインのアタプエルカにあるシマ・デ・ロス・ウエソス遺跡のことです。Sima de los Huesos遺跡は大量の人骨が出ている重要遺跡で、それらの人骨は43万年前頃のものと推定されています(Arsuaga 2014)。Sima de los Huesos遺跡の人骨はネアンデルタール人の人骨と共通点が認められることから、Sima de los Huesos遺跡の人々は初期のネアンデルタール人ではないか、あるいは初期のネアンデルタール人と近縁な集団ではないかという見解が出ていました(Arsuaga 2014)。そのような見解を裏づけるように、Sima de los Huesos遺跡の人々の核DNAは、現生人類とデニソワ人よりネアンデルタール人に近いことが明らかになりました(Meyer 2016)。しかし、不思議なことに、Sima de los Huesos遺跡の人々のミトコンドリアDNAを調べると、ネアンデルタール人よりデニソワ人に近いという結果が出るのです(Meyer 2013、Meyer 2016、Posth 2017)。核DNAを調べると、Sima de los Huesos遺跡の人々はネアンデルタール人に近いが、ミトコンドリアDNAを調べると、Sima de los Huesos遺跡の人々はデニソワ人に近いというわけです。
なぜ核DNAのデータとミトコンドリアDNAのデータが食い違うのでしょうか。このような食い違いがあることがわかったのは最近なので、まだ定説はありません。おそらく、実際の歴史が図2のように単純だったら、核DNAを調べても、ミトコンドリアDNAを調べても、図2のような構図が浮かび上がってきたでしょう。そうならないのは、現生人類とネアンデルタール人とデニソワ人が図2のような枝分かれを起こしたが、枝分かれ後もある程度交わっていたためかもしれません。Posth氏らは、現生人類あるいは現生人類に極めて近い種のミトコンドリアDNAがネアンデルタール人に入って広まった可能性を検討していますが、確かにこれは検討しなければならない問題です。仮にPosth氏らの考える通りであれば、「ミトコンドリアDNAを調べると、現生人類とネアンデルタール人が近く、核DNAを調べると、ネアンデルタール人とデニソワ人が近い」という一見奇妙な状況にも説明がつきます。
※最初に発見されたデニソワ人は女性であり、Y染色体DNAに焦点が当たりませんでしたが、最近になって現生人類、ネアンデルタール人、デニソワ人のY染色体DNAに焦点を当てた研究が発表されました(Petr 2020)。Y染色体DNAを調べても、ミトコンドリアDNAを調べた時と同じように、現生人類とネアンデルタール人が近いという結果が出ています。混迷が深まりそうです。
少し前までは、現生人類の始まりを15~20万年前頃と考えるのが普通でした。東アフリカ(エチオピア)で現生人類のものと判断され、その頃のものと推定される人骨が知られていました(White 2003、McDougall 2005)。
しかし、北アフリカ(モロッコ)で発見された現生人類によく似た何人かの人骨が30万年前頃のものと推定され、流れが変わってきました(Hublin 2017、Richter 2017、日本語のニュース記事はこちら)。
先ほどの図3を見てください。「現生人類」と「ネアンデルタール人とデニソワ人の共通祖先」に枝分かれした箇所があります。Sima de los Huesos遺跡の人々が生きていたのが43万年前頃ですから、「現生人類」と「ネアンデルタール人とデニソワ人の共通祖先」に枝分かれした箇所はもっと前です。現生人類、ネアンデルタール人、デニソワ人、そしてSima de los Huesos遺跡の人々の核DNAを分析したMeyer氏らは、「現生人類」と「ネアンデルタール人とデニソワ人の共通祖先」に枝分かれしたのは55~76.5万年前頃と見積もっています(Meyer 2016)。
現生人類がネアンデルタール人とデニソワ人の共通祖先と枝分かれしたのが55~76.5万年前頃、そして人骨によって確認できる現生人類の始まりが15~20万年前頃ということで、ものすごいギャップがありました。このギャップをいくらか埋めそうなのが、前述の北アフリカ(モロッコ)で発見された30万年前頃の人骨です。この発見によって、現生人類が15~20万年前よりもっと前からアフリカ全体に広がっていた可能性、いやそれどころか、アフリカから出ていた可能性すら出てきました。
6万年前頃にアフリカから出た現生人類が爆発的に拡散して現在に至っています。しかし、イスラエルのSkhul(スクール)やQafzeh(カフゼー)の人骨のように、現生人類がもっと前にアフリカから出ていたことがわずかながら知られています(Shea 2008)。ギリシャ南部のApidima Cave(アピディマ洞窟)で見つかった人骨も加えられそうな気配です(Harvati 2019)。6万年前より前にアフリカから出た現生人類はどうなったのか、少し生き残ったのか、完全に死に絶えたのかという問題は、人類学における争点の一つになっています。
系統的にデニソワ人に近いと考えられるネアンデルタール人が、現生人類に近いミトコンドリアDNAとY染色体DNAを示しているのは、注目に値します。6万年前より前にアフリカから出た現生人類は、他の種を消し去ってしまうような圧倒的な存在ではなく、他の種と混ざり合っていたのかもしれません。
次回の記事で詳しくお話ししますが、現生人類のDNAには、ネアンデルタール人から受け継いだ部分だけでなく、デニソワ人から受け継いだ部分もあります。6万年前頃にアフリカから出た現生人類は、ネアンデルタール人とデニソワ人をいくらか取り込みながら、爆発的に拡散していったのです。ここで気になるのが言語の問題です。
6万年前頃にアフリカから出た現生人類の言語が、現代の言語に近い複雑なものだったのか、それとも現代の言語よりはるかに単純なものだったのかということについては、大いに考察の余地があります。しかし、現生人類はネアンデルタール人とデニソワ人と交わっていたわけですから、ネアンデルタール人とデニソワ人が全く言葉を話さなかったとは考えづらいです。現生人類には自分の言葉があり、ネアンデルタール人とデニソワ人には彼らの言葉があった可能性が高いです。現生人類が言葉を持ち、ネアンデルタール人が言葉を持ち、デニソワ人が言葉を持っていたとすると、これらの三者の共通祖先が言葉を持っていた可能性も少なくありません。ちなみに、人類の脳の進化を振り返ると、アウストラロピテクスの頃には脳容量がチンパンジーと同じくらいだったのに、ホモ・ハビリスの頃から脳容量が急増しています(ホモ・ハビリスはラテン語で「能力のある人」(上手な人、巧みな人、器用な人)という意味です)(図はBretas 2020より引用)。
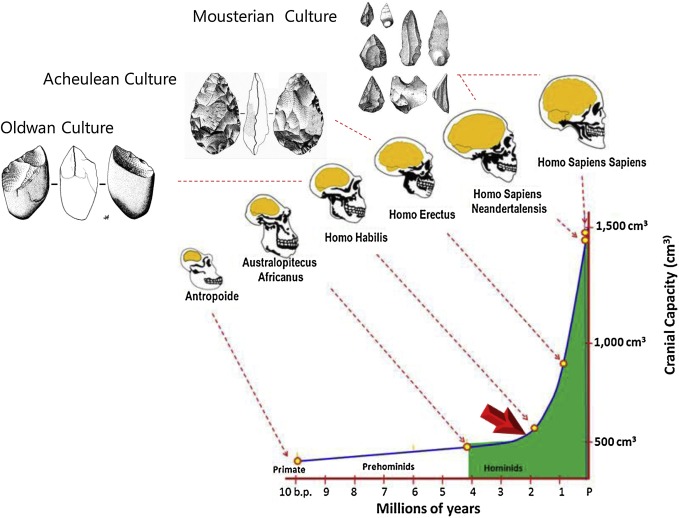
言語の問題は、言うまでもなく、思考と密接な関係にあります。思考が従来と比べて全然発達していないのに、言語がどんどん発達していくというのは、考えづらいです。逆に、思考がどんどん発達しているのに、その伝達手段である言語が全然発達しないというのも、考えづらいです。ホモ・ハビリスの頃から始まる脳容量の急増と言語の形成の間に深い関係があるのは間違いないでしょう。
仮に、現生人類より早くにユーラシアにいたネアンデルタール人とデニソワ人が、さらに早くにユーラシアにいたホモ・エレクトス(北京原人やジャワ原人など)と交わっていたことがわかったら、ホモ・エレクトスが言葉を持っていた可能性も高まります。実際、ネアンデルタール人とデニソワ人がさらに古いタイプの人類と交わっていたかもしれないことを示唆する研究も出てきています(Rogers 2020)。言語が生まれ始めたのは、とても古い時代なのかもしれません。
参考文献
Arsuaga J. L. et al. 2014. Neandertal roots: Cranial and chronological evidence from Sima de los Huesos. Science 344(6190): 1358-1363.
Bretas R. V. et al. 2020. Phase transitions of brain evolution that produced human language and beyond. Neuroscience Research 161: 1-7.
Harvati K. et al. 2019. Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia. Nature 571(7766): 500-504.
Hublin J. J. et al. 2017. New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature 546(7657): 289-292.
Krause J. et al. 2010. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature 464(7290): 894-897.
McDougall I. et al. 2005. Stratigraphic placement and age of modern humans from Kibish, Ethiopia. Nature 433(7027): 733-736.
Meyer M. et al. 2013. A mitochondrial genome sequence of a hominin from Sima de los Huesos. Nature 505(7483): 403-406.
Meyer M. et al. 2016. Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins. Nature 531(7595): 504-507.
Petr M. et al. 2020. The evolutionary history of Neanderthal and Denisovan Y chromosomes. Science 369(6511): 1653-1656.
Posth C. et al. 2017. Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals. Nature Communications 8(1): 16046.
Reich D. et al. 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468(7327): 1053-1060.
Richter D. et al. 2017. The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Nature 546(7657): 293-296.
Rogers A. R. et al. 2020. Neanderthal-Denisovan ancestors interbred with a distantly related hominin. Science Advances 6(8): eaay5483.
Shea J. J. 2008. Transitions or turnovers? Climatically-forced extinctions of Homo sapiens and Neanderthals in the East Mediterranean Levant. Quaternary Science Reviews 27(23-24): 2253-2270.
White T. D. et al. 2003. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423(6941): 742-747.