
前回の記事で示したWen 2016の図を再び掲げましょう。
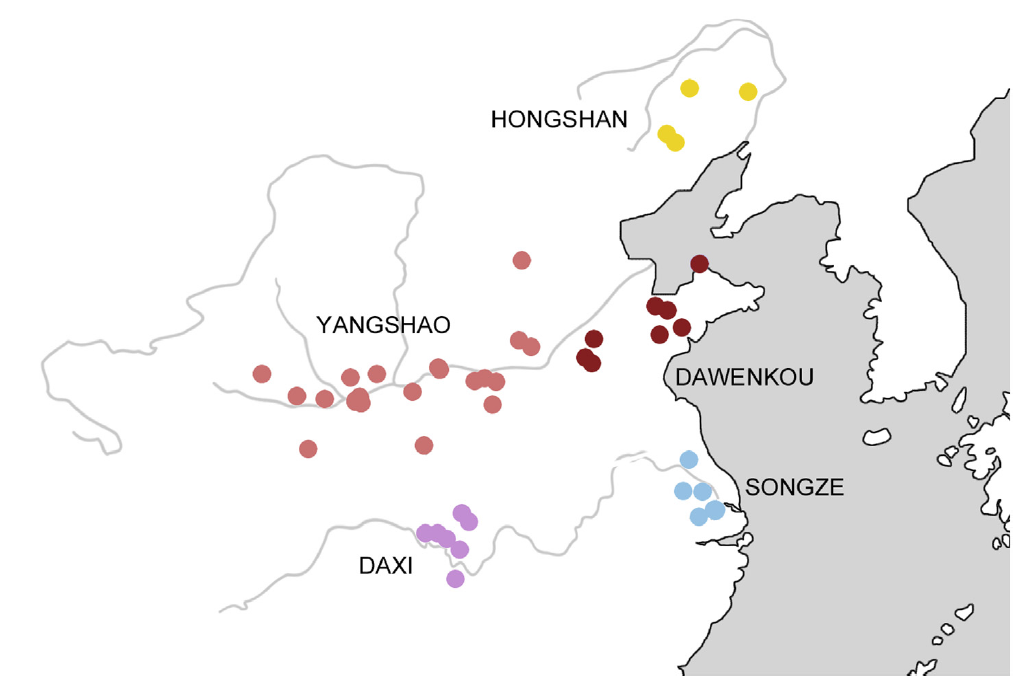
6000年前頃の東アジアの様子です。もうこの頃には本格的な農耕が行われています。北部でアワとキビが栽培され、南部でイネが栽培されたと単純に言われてしまうことが多いですが、北部と南部の間の様相は複雑です。特に、前回の記事で紹介した淮河(わいが)(上の地図には記されていません)は、ちょうど黄河と長江の間にあるので要注意です。
最近の考古学調査により、黄河下流域と長江下流域の間に位置する淮河流域では、8500年前頃からイネの栽培が行われていたことがわかっています(Luo 2016、2019)。稲作発祥の地、つまりイネの栽培が始まったのは、長江下流域~中流域ですが、イネの栽培はかなり早くから北に広がり始めていたということです。
しかし、アワとキビより高温を必要とするイネの栽培は、そうやすやすと北上できたわけではなかったようです。淮河流域よりさらに北に位置する山東省では、苦戦の跡が窺えます。すでにお話ししたように、山東省でイネの栽培が本格的に始まるのは、山東龍山文化の時代(4600~3900年前頃)からです。上の8500年前とこの4600年前では、大きな隔たりがあります。ただし、山東省では山東龍山文化より前にイネの栽培が行われた跡も点々と見つかっており、イネを栽培しようとする試みが繰り返されていたことは窺えます(Luan 2005)。このような試みは決して無駄ではなく、従来のタイプよりいくらか低温に強いタイプの誕生につながったと見られます。
まとめると、こうなります。長江下流域~中流域で始まったイネの栽培は、かなり早い時期に淮河流域に広がった。しかし、淮河流域に広がったイネの栽培は、簡単には山東省に広がることができず、長い苦戦の後で山東省に広がった。
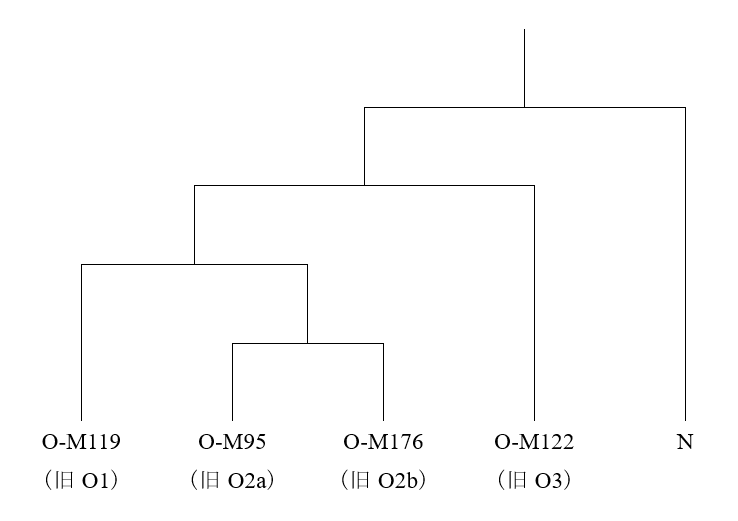
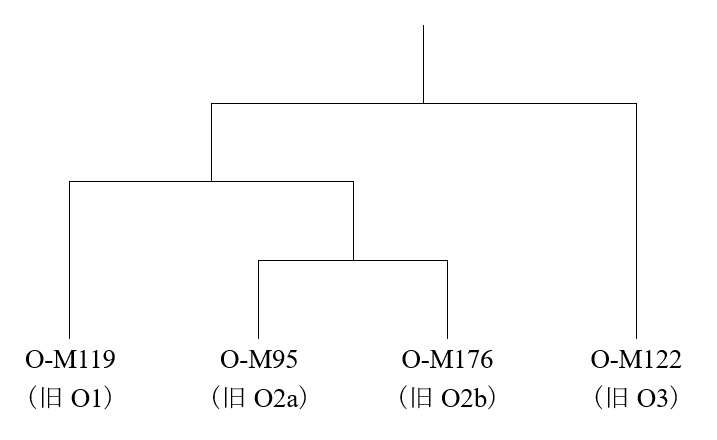
淮河流域でイネを栽培し、その栽培を山東省に広げようと試みていた人間集団はどのような人間集団だったのかということは、大いに考える必要があります。ここで参考になるのが、古代の長江流域のDNAのデータです(Li 2007)。長江下流域(上の地図のSONGZEのあたり)ではY染色体DNAのO-M119が支配的で、長江中流域と下流域の間(DAXIとSONGZEの間のあたり)ではO-M95が支配的で、長江中流域(DAXIのあたり)では複雑だがO-M122が一番多かったことがわかっています。いずれも稲作が盛んに行われていた地域ですが、これらの地域は人間のDNAに関してはそれぞれ明らかに異なる傾向を示しています。南の長江流域でイネを栽培していた人間集団でO-M119とO-M95が支配的だったからといって、北の淮河流域でイネを栽培していた人間集団でもO-M119またはO-M95が支配的だったとは限りません。
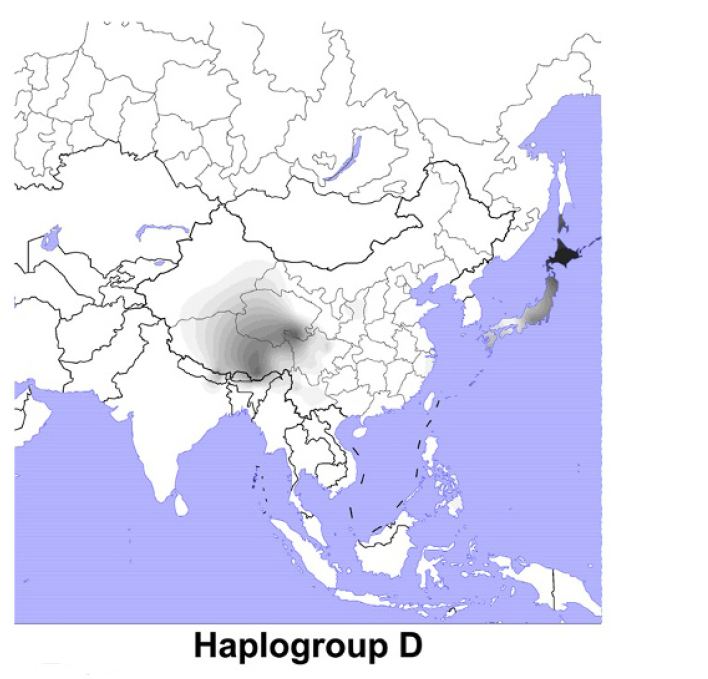
長江流域で行われていたイネの栽培が山東省に伝わり、山東省で行われていたイネの栽培が遼東半島に伝わり、山東省と遼東半島の両方で行われていたイネの栽培が朝鮮半島に伝わり、朝鮮半島で行われていたイネの栽培が日本列島に伝わったと考えられることはお話ししました(最近の考古学のちょっと危ない傾向、遼東半島の稲作をめぐる問題を参照)。しかし、その日本列島では、O-M119とO-M95はわずかしか見られず、代わりに朝鮮半島と共通のO-M176が大きな位置を占めているのです(日本人のY染色体ハプログループOの研究、人と稲作と言語の広がりは必ずしも一致しないを参照)。
前回の記事で説明したように、古代の黄河下流域(DAWENKOUのあたり)のY染色体DNAのデータはまだ発表されていませんが、淮河流域でイネの栽培が行われ、それが山東省に広がり、さらに朝鮮半島に渡る過程において、大きな役割を担ったのは、O-M176の集団であった可能性が高いです。重要なのは、長江流域で行われていたイネの栽培が非常に早い時期(8500年前頃)に淮河流域に伝わっていたという点です。
東アジアの農耕の起源、とても時間がかかる革命、二つの重要な概念の記事で指摘しましたが、従来の歴史研究では以下の二つの人間集団がしっかり区別されてきませんでした(栽培とはなにかということについては、上記の記事で論じたので、そちらを参照してください)。
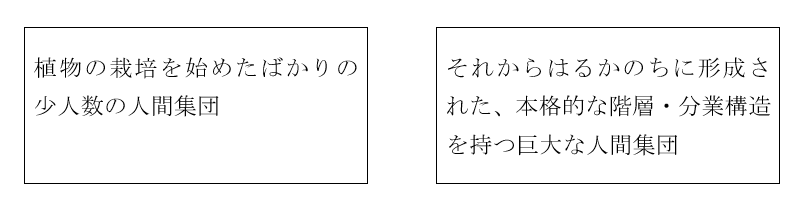
左の人間集団が存在していた時代から右の人間集団が存在する時代への変化は連続的なので、「農耕の起源」と題して一気に語りたくなる気持ちもわからなくはありませんが、やはり歴史を説明するのであれば、最初の段階である左の人間集団と最後の段階である右の人間集団の違いは十分に意識しておく必要があります。だれかが家の庭で行う栽培行為と現代の巨大社会の営みを同じようには語れないでしょう。
特にDNAや言語の問題を考える際には、左の人間集団と右の人間集団の区別に注意しなければなりません。右の人間集団であればあるほど、有無を言わさず、まわりのDNA/言語を消し去り、自分のDNA/言語に置き換えてしまう可能性が高く、左の人間集団であればあるほど、その可能性が低いからです。その可能性が低いというのは、植物の栽培行為自体は広がっていくが、まわりのDNA/言語には大した影響を及ぼさないことも十分ありうるということです。植物の栽培行為自体は、少数の人間を介して伝わることが十分に可能です。
先ほど述べたように、重要なのは、長江流域で行われていたイネの栽培が非常に早い時期(8500年前頃)に淮河流域に伝わっていたという点です。長江流域のO-M119とO-M95の集団が行っていたイネの栽培が淮河流域のO-M176の集団に伝わった可能性が高いです。O-M119とO-M95がO-M176を大きく置き換えることはなかったということです。朝鮮半島と日本列島にO-M176が多く見られ、O-M119とO-M95がわずかしか見られないことから、そのように推測されます(朝鮮半島のデータについてはKim 2010、日本列島のデータについてはNonaka 2007を参照)。
ここまでは、ほぼ朝鮮半島と日本列島に限られているO-M176に焦点を当ててきましたが、朝鮮半島と日本列島にはO-M176だけでなくO-M122も多いので、ここからは、O-M176とO-M122の双方に注目しながら話を進めましょう。
古代中国の青銅器はどこから来たのか
古代中国の青銅器は非常に高度で、古代中国の青銅器は一体どこから来たのだろうと議論されてきました。青銅とは、銅にスズがいくらか混ざったものです。現在では、世界中の考古学データが蓄積し、銅(あるいは金属一般)の利用が中東から世界に広まっていったことが明らかになっています。以下の図は、Roberts 2009からの引用です。
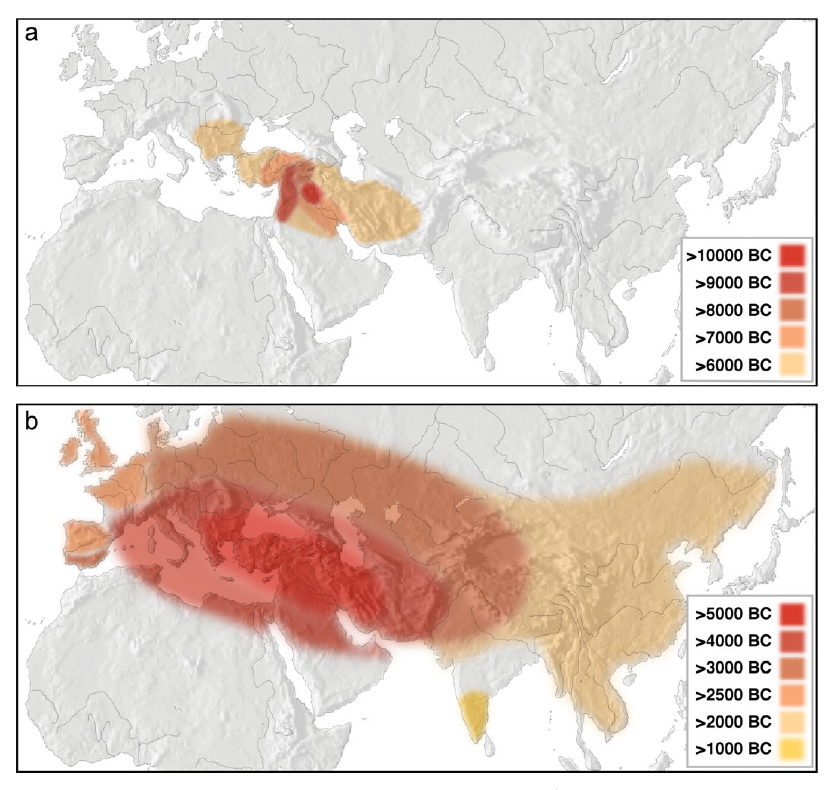
BCは紀元前という意味です。上は、人類が自然界にほぼ単体で存在する銅(とても少ないです)または銅を含む銅鉱石を集めていた時代です。下は、人類が銅の製錬(銅鉱石からお目当ての銅を取り出す作業)を行っている時代です。
古代中国の青銅器の起源探しは続けられていますが、細かな時期やルートに検討の余地は残っているものの(Mei 2015)、銅(あるいは金属一般)の利用が中東から始まったという事実は揺らぎそうにありません。
ただし、夏と推定される二里頭文化の時代とその後の殷の時代に青銅器の製作が著しく高度化したのも事実です。二里頭文化の前から中国に銅器は点々と見られましたが、二里頭文化の時代に大きな発達を遂げ、殷の時代にさらに大きな発達を遂げました(Xu 2021)。殷は夏を滅ぼした後も二里頭遺跡とそのすぐそばにとどまり、筆者はなにをしているのかと疑問に思っていましたが、夏が持っていた施設や技術者を活用していたようです(Xu 2021)。このあたりの合理的な判断がのちの殷の繁栄にうまくつながったと見られます。
古代中国の高度な青銅器の製作は中国で発達したものだが、最も素朴な銅器作りは中国の外からやって来たものだということです。
金属の利用(当初は装飾のためだったと考えられています(Roberts 2009))が始まり、広まったことは人類の歴史において大きな出来事でしたが、だからといって単純に中東のDNA/言語が世界を席巻したわけではありません。ある文明・文化的特徴の広がりとDNA/言語の広がりは必ずしも同一視できません。
確かに、農耕の起源は、本格的な階層・分業構造を持つ巨大な人間社会の誕生を可能にしたという意味で、金属の利用の始まり以上の大きな出来事でしょう。しかし、農耕の起源の最初の場面に目を向ければ、それは素朴な栽培行為の始まりであり、その素朴な栽培行為は、素朴な銅器作りと同様に、必ずしも伝わる先々のDNA/言語に大きな影響を及ぼすものではなかったと考えられるのです。
ちなみに、モンゴルの伝統的な遊牧民としての生活スタイルは、農耕と対照的ですが、モンゴルのあの生活スタイルは、もともと東アジアにあったものではなく、西側からもたらされたものです。しかし、モンゴル人に西側からのDNAがどのくらい入っているかというと、驚くほど少ないのです(Jeong 2020)。モンゴル語も、遊牧民の生活スタイルとともに西側からもたらされたとはとても考えられないものです。ある文明・文化的特徴の広がりをDNA/言語の広がりに単純に結びつけることはできないのです。
参考文献
英語
Jeong C. et al. 2020. A dynamic 6,000-year genetic history of Eurasia’s Eastern Steppe. Cell 183(4): 890-904.
Kim H. et al. 2010. Y chromosome homogeneity in the Korean population. International Journal of Legal Medicine 124(6): 653-657.
Li H. et al. 2007. Y chromosomes of prehistoric people along the Yangtze River. Human Genetics 122(3-4): 383–388.
Luo W. et al. 2016. Phytolith records of rice agriculture during the Middle Neolithic in the middle reaches of Huai River region, China. Quaternary International 426: 133-140.
Luo W. et al. 2019. Phytoliths reveal the earliest interplay of rice and broomcorn millet at the site of Shuangdun (ca. 7.3–6.8 ka BP) in the middle Huai River valley, China. Journal of Archaeological Science 102: 26-34.
Mei J. et al. 2015. Archaeometallurgical studies in China: Some recent developments and challenging issues. Journal of Archaeological Science 56: 221-232.
Nonaka I. et al. 2007. Y-chromosomal binary haplogroups in the Japanese population and their relationship to 16 Y-STR polymorphisms. Annals of Human Genetics 71(4): 480-495.
Roberts B. W. et al. 2009. Development of metallurgy in Eurasia. Antiquity 83(322): 1012-1022.
Wen S. et al. 2016. Y-chromosome-based genetic pattern in East Asia affected by Neolithic transition. Quaternary International 426: 50-55.
その他の言語
Luan F. 2005. 海岱地区史前时期稻作农业的产生、发展和扩散. 文史哲 6: 41-47.(中国語)
Xu C. 2021. 陶寺、石峁之争与二里头的崛起:寻找中国历史上第一个王朝——夏. 国家人文历史 15: 30-37.(中国語)