日本語の起源と歴史に興味を持つすべての方へ
こんにちは。金平譲司と申します。ここに「日本語の意外な歴史」と題するブログを立ち上げました。
このブログは、日本語ならびに日本語と深い関係を持つ言語の歴史を解明するものです。言語学者だけでなく、他の分野の専門家や一般の方々も読者として想定しています。
謎に包まれてきた日本語の起源
日本語はどこから来たのかという問題は、ずいぶん前から様々な学者によって論じられてきましたが、決定的な根拠が見つからず、大いなる謎になってしまった感があります。しかしながら、筆者の研究によってようやくその全貌が明らかになってきたので、皆さんにお話ししようと思い立ちました。
日本語は、朝鮮語、ツングース諸語(エヴェンキ語、満州語など)、モンゴル諸語(モンゴル語、ブリヤート語など)、テュルク諸語(トルコ語、中央アジアの言語など)と近い関係にあるのではないか、あるいはオーストロネシア語族(台湾、フィリピン、インドネシア、マレーシア、オセアニアなどの言語から成る言語群)と近い関係にあるのではないかというのが従来の大方の予想でしたが、これらの予想はポイントを外しています。
中国語を見て全く違うと感じた日本人が、日本語は北方の言語と関係があるのではないか、南方の言語と関係があるのではないかと考えたのは、至極当然のことで、北方の言語と南方の言語に視線を注ぐこと自体は間違っていません。問題なのは、北方のごく一部の言語と南方のごく一部の言語に関心が偏ってしまったことです。
上記の言語のうちで、朝鮮語、ツングース諸語、モンゴル諸語、テュルク諸語は、日本語によく似た文法構造を持つことから、日本語に近縁な言語ではないかと盛んに注目されてきました。同時に、ツングース諸語、モンゴル諸語、テュルク諸語は、互いに特に近い関係にあるとみなされ、いわゆる「アルタイ語族」という名でひとまとめにされることがしばしばありました。日本語の起源をめぐる議論は、このような潮流に飲まれていきました。
しかしながら、筆者がこれから明らかにしていく歴史の真相は、かなり違います。日本語は、朝鮮語、ツングース諸語、モンゴル諸語、テュルク諸語と無関係ではないが、別の言語群ともっと近い関係を持っているようなのです。
実を言うと、筆者は日本語やその他の言語の歴史に興味を持つ人間ではありませんでした。筆者は若い頃にフィンランドのヘルシンキ大学で一般言語学や様々な欧州言語を学んでいましたが、その頃の筆者の興味は言語と思考の関係や外国語の学習理論などで、もっぱら現代の言語に関心が向いていました。歴史言語学の講義もありましたが、特に気に留めていませんでした。
筆者が言語の歴史について真剣に考えるようになったきっかけは、ロシアの北極地方で少数民族によって話されているサモエード諸語との出会いでした。サモエード諸語は、フィンランド語やハンガリー語と類縁関係にある言語です。フィンランド語とハンガリー語はヨーロッパの中では異色の存在で、北極地方の少数民族の言語と類縁関係を持っています。フィンランド語、ハンガリー語、サモエード諸語などから成る言語群は、「ウラル語族」と呼ばれます。
言語学者が使う「語族」という用語について若干説明しておきます。私たちが万葉集や源氏物語の言葉を見ると、「読みにくいな」と感じたり、「なにを言っているのかわからないな」と感じたりします。言語は時代とともに少しずつ変化しています。言語は単に変化するだけでなく、分化もします。ある程度広い範囲で話されている言語には、地域差が生じてきます。
この地域ごとに少しずつ異なる言葉が方言です。しかし、これらの方言が地理的に隔たってさらに長い年数が経過すると、最初は小さかった方言同士の差が大きくなっていき、やがて意思疎通ができないほどになります。
あまりに違いが大きくなれば、もう方言ではなく、別々の言語と言ったほうがふさわしくなります。一律の学校教育やマスメディアが発達していない時代には、この傾向は顕著です。ある言語が別々の言語に分化するのです。分化してできた言語がさらに分化することもあります。言語学では、おおもとの言語と分化してできた諸言語をまとめて「語族」といいます。世界で最もよく知られている語族は、インド・ヨーロッパ語族と呼ばれる語族で、英語、ドイツ語、フランス語、ロシア語などはこの語族に属します。例えるなら、イヌ、オオカミ、キツネ、タヌキが共通祖先を持っているように、英語、ドイツ語、フランス語、ロシア語は共通祖先を持っているということです。
日本語とウラル語族
英語などが属するインド・ヨーロッパ語族は巨大な言語群ですが、フィンランド語やハンガリー語が属するウラル語族はこじんまりとした言語群です。ウラル語族の言語は、ロシアの北極地方から北欧・東欧にかけて分布しています。地理的に遠く離れているので、ウラル語族の言語は一見したところ東アジアの言語、特に日本語とはなんの関係もないように見えますが、実はここに大きな盲点があります。日本語の歴史を考えるうえで大変重要になるので、ウラル語族の話を続けます。以下にウラル語族の内部構造を簡単に示します。
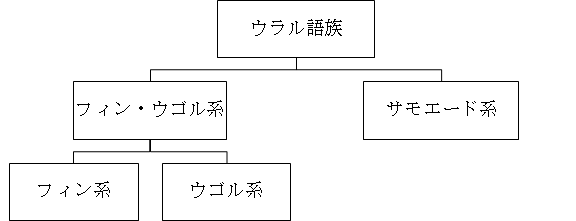
ウラル語族の言語を研究する学者の間に意見の相違がないわけではありませんが、上の図は従来広く受け入れられてきた見方です。ウラル語族の言語は、まずフィン・ウゴル系とサモエード系に分かれ、フィン・ウゴル系はそこからさらにフィン系とウゴル系に分かれます。フィンランド語はフィン系に属し、ハンガリー語はウゴル系に属します。サモエード系の言語は、ロシアの北極地方に住む少数民族によって話されています。現在残っているサモエード系の言語はネネツ語、エネツ語、ガナサン語、セリクプ語の四つのみで、特に後の三つは消滅の危機にあります。
サモエード系の言語は、フィンランド語やハンガリー語と同じウラル語族の言語ですが、フィンランド語やハンガリー語とは文法面でも語彙面でも著しく異なっています。同じ言語から分かれた言語同士でも、別々の道を歩み始め、何千年も経過すれば、似ても似つかない言語になってしまいます。特に、サモエード系の言語が辿った運命とフィンランド語・ハンガリー語が辿った運命は対照的です。サモエード系の言語は、北極地方にとどまり、他の言語との接触が比較的少なかったために、昔の姿をよく残しています。それに対して、フィンランド語とハンガリー語は、有力な言語がひしめくヨーロッパに入り込み、大きく姿を変えました。サモエード系の言語は、いわば「生きた化石」です。人類の歴史を解明するうえで、大変重要な言語です。サモエード系の言語との出会いは、筆者にとってショッキングな出来事でした。これ以降、筆者は言語の歴史について本格的に研究し始めることになります。
筆者が初めてサモエード系の言語を見た時には、「文法面ではモンゴル語やツングース諸語に似ているな」という第一印象を受けました。しかし、よく調べると、「あれっ、語彙面では日本語に似ているな」という第二印象を受けました。少なくとも言語の根幹をなす基礎語彙に関しては、モンゴル語やツングース諸語より、ウラル語族のサモエード系の言語のほうが日本語に近いと思いました。なんとも不思議な感じがしました。なんで日本の近くで話されているモンゴル語やツングース諸語より、北極地方で話されているウラル語族のサモエード系の言語のほうが日本語に近いんだろうと考え始めました。様々な言語を見てきましたが、サモエード系の言語には今までにない特別なものを感じました。なにか重大な秘密が隠されている予感がしました。
フィンランド語とハンガリー語だけを見ていた時は気づかなかったのですが、サモエード系の言語を介しながらフィンランド語とハンガリー語を見てみると、やはりフィンランド語とハンガリー語にも日本語との共通語彙があります。日本語の中にある、ウラル語族と共通している語彙、そしてウラル語族と共通していない語彙を見分けていくうちに、二つの疑問が頭に浮かんできました。一つ目の疑問は、日本語の祖先とウラル語族の言語の祖先の接点は地理的にどの辺にあったのだろうという疑問です。二つ目の疑問は、日本語の中にある、ウラル語族と共通していない語彙はどこから来たのだろうという疑問です。日本語の中には、ウラル語族と共通している語彙も多いですが、共通していない語彙も多いのです。
東アジアには黄河文明とは違う文明が存在した
ウラル語族の各言語の語彙を研究するうちに、ウラル語族が日本語だけでなく、モンゴル語、ツングース諸語、朝鮮語、さらには中国語にもなんらかの形で関係していることが明らかになってきたので、ウラル語族の言語と東アジア・東南アジアの言語の大々的な比較研究を開始しました。着実かつ合理的に歴史を解明するため、考古学および生物学の最新の研究成果を適宜参照しました。考古学も生物学も近年めざましい発展を遂げており、数々の重要な発見がありました。
かつては、メソポタミア文明、エジプト文明、インダス文明、そして東アジアの黄河文明が並べられ、世界四大文明と呼ばれていました。ところが、その後の発見によって、東アジアには黄河文明のほかに二つの大きな文明が存在したことがわかってきました(このテーマを包括的に扱った書籍はいくつかありますが、考察の広さ・深さの点でShelach-Lavi 2015が優れています)。
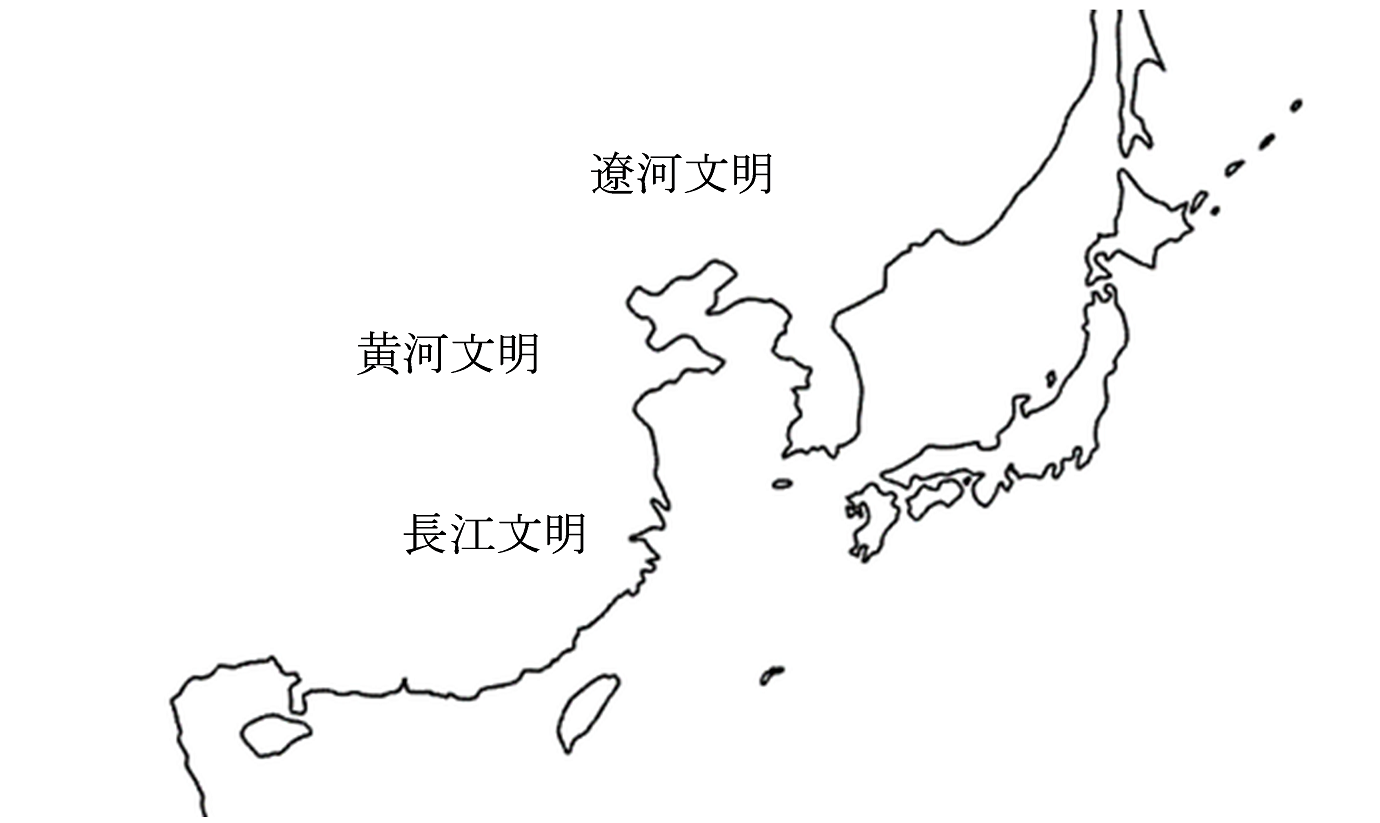
その二つの大きな文明とは、長江文明と遼河文明(りょうがぶんめい)です。日本列島で縄文時代が進行する間に、大陸側はこのようになっていたのです。黄河文明と長江文明に比べて、遼河文明は知名度が高くないかもしれません。しかし、遼河文明は、日本語の歴史を解明するうえで重要な鍵を握っているようなのです。
生物学が発達し、人間のDNA配列が調べられるようになりました。DNA配列は、正確には「DNAの塩基配列」といい、アデニンA、チミンT、グアニンG、シトシンCという四種類の物質が作る列のことです。最近では、生きている人間のDNA配列だけでなく、はるか昔に生きていた人間のDNA配列も調べられるようになってきました。大変興味深いことに、遼河文明が栄えていた頃に遼河流域で暮らしていた人々のDNA配列を調べた研究があります(Cui 2013)。
人間は父親と母親の間に生まれるので、子のDNA配列が父親のDNA配列と100パーセント一致することはなく、子のDNA配列が母親のDNA配列と100パーセント一致することもありません。しかし、父親から息子に代々不変的に受け継がれていく部分(Y染色体DNA)と、母親から娘に代々不変的に受け継がれていく部分(ミトコンドリアDNA)があります。代々不変的に受け継がれていく部分と書きましたが、この部分にも時に突然変異が起きます。つまり、その部分のDNA配列のある箇所が変化するのです。変化していないY染色体DNA配列を持つ男性がそれを息子に伝える一方で、変化したY染色体DNA配列を持つ男性がそれを息子に伝えるということが起き始めます。同様に、変化していないミトコンドリアDNA配列を持つ女性がそれを娘に伝える一方で、変化したミトコンドリアDNA配列を持つ女性がそれを娘に伝えるということが起き始めます。こうして、時々起きる突然変異のために、Y染色体DNAのバリエーション、ミトコンドリアDNAのバリエーションができてきます。人類の歴史を研究する学者は、このY染色体DNAのバリエーション、ミトコンドリアDNAのバリエーションに注目するのです。
先ほど述べた遼河流域の人々のDNA研究は、Y染色体DNAのバリエーション(例えば、C系統か、D系統か、N系統か、O系統か)を調べたものです。その結果はどうだったでしょうか。古代の人々の研究なのでサンプル数は限られていますが、それでも大まかな傾向は十分に捉えられています。遼河文明が栄えていた頃の遼河流域では、当初はN系統が圧倒的に優勢だったが、次第にO系統とC系統が増え(つまり他の地域から人々が流入してきたということ)、N系統はめっきり少なくなってしまったようです。現在の日本、朝鮮半島、中国では、N系統はほんの少し見られる程度です(Shi 2013)。対照的に、ウラル語族の言語が話されているロシアの北極地方からフィンランド方面にかけてN系統が非常に高い率で観察されています(Rootsi 2007)。
見え始めた日本語の正体
筆者もウラル語族の言語が東アジアの言語と深い関係を持っていることを知った時には大いに驚きましたが、考古学・生物学の発見と照らし合わせると、完全に合致します。日本語がウラル語族の言語と深い関係を持っていることは非常に興味深いですが、もう一つ興味深いことがあります。日本語の中には、ウラル語族と共通している語彙も多いですが、共通していない語彙も多く、ウラル語族とは全く異なる有力な言語群も日本語の形成に大きく関与したようなのです。
ウラル語族の言語と東アジア・東南アジアの言語の大々的な比較研究を行い、様々な紆余曲折はありましたが、漢語流入前の日本語(いわゆる大和言葉)の語彙構成が以下のようになっていることがわかってきました。
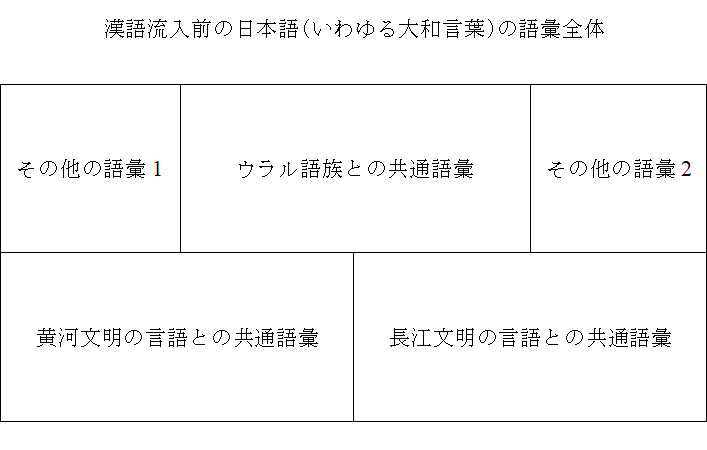
「ウラル語族との共通語彙」も多いですが、「黄河文明の言語との共通語彙」と「長江文明の言語との共通語彙」も多く、この三者で漢語流入前の日本語の語彙の大部分を占めています。
「その他の語彙1」というのは、日本語が大陸にいた時に取り入れた語彙で、「ウラル語族との共通語彙」にも、「黄河文明の言語との共通語彙」にも、「長江文明の言語との共通語彙」にも該当しないものです。
「その他の語彙2」というのは、日本語が縄文時代に日本列島で話されていた言語から取り入れた語彙です。
漢語流入前の日本語の語彙構成の特徴的なところは、なんといっても、語彙の大きな源泉が三つあることです。三つの有力な言語勢力が交わっていたことを窺わせます(遼河文明と黄河文明と長江文明の位置を思い出してください)。
「日本語の意外な歴史」では、ウラル語族との共通語彙、黄河文明の言語との共通語彙、長江文明の言語との共通語彙、その他の語彙1、その他の語彙2、いずれも詳しく扱っていきます。
では、日本語およびその他の言語の歴史を研究するための準備に取りかかりましょう。
外国語の単語の表記について
英語と同じようなアルファベットを使用している言語では、それをそのまま記します。言語学者が諸言語の発音を記述するのに使う国際音声記号(IPA)というのがありますが、音韻論の専門家でない限り、多くが見慣れない記号です。そのため、本ブログではIPAの使用はできるだけ控えます。特に朝鮮語は、IPAを用いて記すと複雑になるため、市販されている初心者向けの韓国語の文法書で採用されている書き方にならいました。一般の読者にとって見慣れない記号を用いる場合には、補助としてのカタカナ表記を付け加えます。慣習を考慮し、ヤ行の子音は基本的に、北方の言語(ウラル語族の言語など)では「j」で表し、南方の言語(中国語、東南アジアの言語)では「y」で表します。古代中国語のアルファベット表記の仕方は、Baxter 2014に従います。
参考文献
Baxter W. H. et al. 2014. Old Chinese: A New Reconstruction. Oxford University Press.
Cui Y. et al. 2013. Y chromosome analysis of prehistoric human populations in the West Liao River Valley, Northeast China. BMC Evolutionary Biology 13: 216.
Rootsi S. et al. 2007. A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe. European Journal of Human Genetics 15: 204-211.
Shelach-Lavi G. 2015. The Archaeology of Early China: From Prehistory to the Han Dynasty. Cambridge University Press.
Shi H. et al. 2013. Genetic evidence of an East Asian origin and paleolithic northward migration of Y-chromosome haplogroup N. PLoS One 8(6): e66102.
かつての日本語の隣人をよく知る
「生意気(なまいき)」とは何か、誰もが違和感を覚える「舐める」と「ナメる」の記事で「意味の干渉」についてお話しし、タイミングがよいので、日本語の隣人の話をします。
「水」のことをmat-、mit-、mut-、met-、mot-のように言ったり、bat-、bit-、but-、bet-、bot-のように言ったり、pat-、pit-、put-、pet-、pot-のように言ったりしていた隣人です。日本語にとっては、単なる隣人ではなく、近縁関係にある隣人です。
これらの隣人をよく知ることは、非常に重要です。日本語が大陸にいた時に、日本語が中国語と朝鮮語に直接接触しなくても、日本語の隣人が中国語と朝鮮語に直接接触した可能性があるからです(無数の小さい言語が存在していた時代には、後者のほうがはるかに可能性が高いです)。
話の都合上、「水」のことをpataのように言っていた隣人を中心に見ていきます。
「水」のことをpataのように言っていた日本語の隣人が、朝鮮語に出会ったら、どうなるでしょうか。pataは、「水」を意味することができなくなって、「海」を意味しようとするかもしれません。「川」を意味しようとするかもしれません。「雨」を意味しようとするかもしれません。

実際、朝鮮語では、「海」のことをpadaと言います。「川」は、古代中国語のkæwng(江)カウンを取り入れて、kaŋカンと言います。「雨」のことは、piと言います。
pada(海)は、日本語の近縁言語から入った可能性が非常に高いです。pi(雨)は、一音節なので、検討の余地が残ります(しかし、日本語のpityapitya(ぴちゃぴちゃ)、ɸitu(漬つ)(未然形ɸita)、ɸitasu(浸す)、ɸitaru(浸る)などの語から、「水」のことをpitaのように言う言語があったことは確実です)。
日本語のbatabata(ばたばた)とbasabasa(ばさばさ)、batyabatya(ばちゃばちゃ)とbasyabasya(ばしゃばしゃ)のような語を見れば、[t]~[tʃ]~[ʃ]~[s]の間で発音変化が頻繁に起きていたこともわかります([t]は「タ、ティ、トゥ、テ、ト」の類、[tʃ]は「チャ、チュ、チョ」の類、[ʃ]は「シャ、シュ、ショ」の類、[s]は「サ、スィ、ス、セ、ソ」の類です)。
そうであるなら、朝鮮語のmasida(飲む)マシダ(daは動詞・形容詞に付く形式的な要素です)も、「水」のことをmat-、mit-、mut-、met-、mot-のように言っていた日本語の近縁言語から入った可能性が高いです。
※朝鮮語にはmit(下)ミ(トゥ)という語もあり、これも絶対に無視できません。「水」→「雨」→「落下、下方向、下」の超頻出パターンです。
上のpada(海)やmasida(飲む)が非常に重要なのは、「水」→「海」という意味変化、「水」→「飲む」という意味変化がダイレクトだからです。「水」が、いくつもの意味変化を経て、「海」を意味するようになった、あるいは、「水」が、いくつもの意味変化を経て、「飲むこと」を意味するようになった、そういう間接的なストーリーは考えられないのです。日本語の近縁言語が直接、朝鮮語に接したということです。
日本語のumi(海)とuna(海)は、ミャオ・ヤオ系言語から入った語で、nomu(飲む)は、タイ系言語から入った語でした。同じように、朝鮮語のpada(海)は、日本語の近縁言語から入った語で、masida(飲む)も、日本語の近縁言語から入った語のようです。これは、順当と言ってよいでしょう。朝鮮語も、多数の言語が消えていく中で生き残った有力な言語です。遼河文明の影響を受けることがなければ、それは不可能だったでしょう。
要するに、日本語の歴史はもちろん、朝鮮語の歴史を知るうえでも、東アジアの歴史を知るうえでも、日本語の近縁言語を知ることは重要だということです(お話しするのはまだ先になりますが、北ユーラシアの壮大な歴史を知るうえでも、大変重要になってきます)。
日本語の近縁言語を知ることの重要性を認識したうえで、「水」のことをpataのように言っていた言語をクローズアップしましょう。
いざ本題へ
先ほどの図をもう一度貼ります。

まずは、海の図から始めましょう。
「水」を意味していた語が、「海」または「波」を意味するようになるのは、よくあるパターンです。しかし、「海」も「波」も人気の行き先なので、すぐに他の語に占められてしまいます。「海」も「波」も意味できない場合は、どうしたらよいでしょうか。
前に、波に揺られての記事で少しお話ししましたが、「揺れること、動くこと」を意味するようになるのです。陸の上に置いた物は、動かないでしょう。しかし、水の上に浮かんでいる物は、どうでしょうか。ふらふらと動きますね(uku(浮く)(未然形uka)、ukabu(浮かぶ)とugoku(動く)、ugomeku(蠢く)を見ると、似ていないでしょうか)。
日本語にɸataraku(働く)という語がありました。現代のhataraku(働く)とはちょっと違っていました。ɸataraku(働く)の使用例を岩波古語辞典(大野1990)から引いてみます。
- 「俄かに弓に箭を番ひて、本の男に差し充て強く引きて、『おのれ働かば射殺してむ』と云へば」(今昔物語集)
- 「死にて六日といふ日の未の時ばかりに、にはかにこの棺働く」(宇治拾遺物語)
上の例は、弓矢を引いて、動いたら殺すぞと言っています。下の例は、死体が入っているはずの棺が動いたと言っています。
これらの例からわかるように、ɸataraku(働く)は、ugoku(動く)と同じ意味でした。
「水」を意味していたpataのような語が、「水」を意味できず、「海」も「波」も意味できず、「動くこと」を意味するようになったのです。
現代の日本語でも、「今、ばたばたしておりまして」などと言いますね。このbatabata(ばたばた)も同じところから来ており、動きまわることを意味しているのです。
zitabata(じたばた)も関係があるでしょう。この語は、「足」を意味するsitaと「動くこと」を意味するpataがくっついたと見られます。日本語では、昔からiとuの間の発音変化が盛んで(一年ぶりの記事、まずは昔の話題の続きから、ついにベールを脱ぐミャオ・ヤオ語族を参照)、sutasuta(すたすた)という語が残っているので、sitaが「足」を意味することもあったと考えられます。
dotabata(どたばた)も、騒がしく歩くあるいは走ることを表すdotadota(どたどた)があるので、「足」+「動く」でしょう。
ɸataraku(働く)と同様に、ɸatameku(はためく)も、「動くこと」を意味していたはずです。しかし、ɸatameku(はためく)という動詞では、「意味の干渉」が強く起きているように見受けられます(「意味の干渉」については、「生意気(なまいき)」とは何か、誰もが違和感を覚える「舐める」と「ナメる」を参照)。

左のɸataに、右のɸataが干渉してきます。その結果、ɸatameku(はためく)は、かつてのように自由に動きを表すことはできず、旗のような動きしか表せなくなったのです。
ちなみに、ɸata(旗)はどこから来たのでしょうか。
奈良時代の日本語には、ɸata(旗)のほかに、ɸata(機)という語がありました。ɸata(機)は、布を織る機械です。「布を織る」とは、経糸(たていと)と緯糸(よこいと)を組み合わせて布を作ることです(図は藤岡糊付所様のウェブサイトより引用)。

要するに、ɸata(機)とは、糸から布を作る機械です。
ɸata(旗)とɸata(機)には、「糸」が共通しています。
現代人は、ほとんど完成品を買うだけですが、昔の人にとっては、糸から布、布から服に至るプロセスはもっと身近であったと考えられます。そのプロセスを自分で行わないとしても、身近にいるだれかが行うのを見ていたでしょう。糸の状態でも、布の状態でも、服の状態でも、kinu(絹、衣)という語が使われていましたが、これも、全く違和感のないことだったのでしょう。
豊富な糸関連の語彙の中で、ɸataは「糸」を意味しようとしたが、他の語に押され、糸から作られる織物(ɸata(旗))と、糸から織物を作る機械(ɸata(機))を意味するようになったと見られます。
「水」を意味していた語が「糸」を意味するようになるのは、頻出パターンです。「水」を意味していた語が、「(水と陸の)境」を意味するようになり、「境」を意味していた語が、「線状のもの」を意味するようになるのです。「水」を意味していたpataのような語も、このパターンをたどったと見られます。
ミャオ・ヤオ系言語で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語は、amu(編む)(未然形ama)、ami(網)になったと考えられます。タイ系言語で「水」を意味したnam、nim、num、nem、nomのような語は、日本語の糸関連の語彙を見る限り、naɸa(縄)、naɸu(綯う)(未然形naɸa)(糸などをねじり合わせる作業)、nuɸu(縫ふ)などになったのではないかと思われます。三省堂時代別国語大辞典上代編(上代語辞典編修委員会1967)が指摘しているように、niɸiname(新嘗)がかつてniɸinami、niɸinaɸe、niɸinaɸiと呼ばれていたことを考えれば、namのmの部分がbになるだけでなく、p(またはɸ)になることもあったと考えられます。一般に、m~b~p(またはɸ)の間は発音変化が起きやすいところなので、そう考えることに無理はありません。むしろ、m~bの間でだけ変化が起きて、b~p(またはɸ)の間では変化が起きないと考えるほうが不自然です。niɸiname(新嘗)というのは、神に新穀を捧げて収穫を感謝し、自らもそれを食べる古来の儀式のことです。
※ミャオ・ヤオ語族から日本語にuna(海)という語が入りましたが、この語も「動くこと」を意味するようになったようです。まずは、uneru(うねる)です。現代でも「海のうねり」と言いますね。水の上に浮かんでいる物がふらふらと動いているところを想像していただければと思いますが、特に向きを変える動きを意味しやすいようです。uneune(うねうね)がまさにこれです。
ややわかりにくいのが、unagasu(促す)です。英語で「促すこと」をurgeと言ったり、promoteと言ったりします。urgeも、promoteも、昔は今ほど抽象的ではなく、「前に押すこと、前に動かすこと」を意味していました。日本語のunagasu(促す)もこれです。ただ、かつて存在したはずの自動詞のunagu*が完全に消えており、他動詞のunagasu(促す)だけが残っているので、わかりにくくなっています。
次に、川の図に移りましょう。
これは、「水」を意味していた語が、横の部分を意味するようになるパターンです。

「水」を意味していたpataのような語が、横の部分を意味するようになり、日本語のɸata(端、辺)になったことは、すでにお話ししました。そこで終わらず、ɸata(畑)/ ɸatake(畑)(古形ɸataka*)やɸada(肌)/ɸadaka(裸)にもなったという、びっくりする話もしました(前回の記事を参照)。
ここでは、それ以外の話をしましょう。
「水」を意味したpataのような語は、奈良時代の日本語のɸata(端)とɸasi(端)からわかるように、あまり変わっていない形で日本語に残ることもあれば、大きく変わった形で日本語に残ることもありました。たどってきた経緯が少し違うということです。ɸati*やɸasa*のような形もあったでしょう。
「水」を意味していた語は、「陸地」を意味するようになることがありますが、「水と陸の境」を意味するようになることもあります(先ほどお話しした、「糸」を意味しようとしたがそれができなかったɸata(旗)とɸata(機)も、この後者のケースです)。

ややこしいことに、この「水と陸の境」を意味する語が、「間」を意味するようになることがあるのです。
実際、奈良時代の日本語には、ɸasi(端)という語のほかに、もう消滅しかかっていましたが、ɸasi(間)という語がありました。
「端」を意味する語がɸasiで、「間」を意味する語がɸasiだと、さすがに都合が悪いです。だから、ɸasi(間)は消滅しかかっていたのでしょう。消滅しかかったɸasi(間)はどうやら、両岸の間に設置されるɸasi(橋)になったようです。昔の日本人は、現代なら「はしご」や「階段」と呼ぶようなものまで、ɸasi(橋)と呼んでいました。隔たりのある二地点の「間をつなぐもの」という認識だったのでしょう。
ɸasi(間)のほかに、ɸasa*(間)という語もあったと思われます。
ɸasi(端)とɸasi(間)よりはましですが、ɸasi(端)とɸasa*(間)も紛らわしいです(しかも、ɸata(端)もあります)。
ɸasi(間)は、ɸasi(橋)になって生き残りましたが、ɸasa*(間)は、類義語のma(間)を結合し、ɸasama(はさま)として生き残ったようです。ɸasama(はさま)から、ɸasamu(はさむ)、ɸasamaru(はさまる)、さらにɸasami(はさみ)ができ、ɸasama(はさま)自身は、ɸazama(はざま)と濁りました。
ɸasi(間)は、場所を意味するta(konata(こなた)やkanata(かなた)のta)を結合して、「間」という意味を保とうとしたこともあったかと思われます。その名残が、中途半端であることを意味したɸasita(はした)です。「はした金」はもともと、中途半端なお金のことでした。今では、わずかなお金を意味するのが普通でしょう。
ɸasitanasi(はしたなし)も関係がありますが、もっと難しいです。忙しいことを意味するseɸasi(せはし)にnasi(なし)がくっついて強調されたのがseɸasinasi(せはしなし)ですが、それと同様に、中途半端であることを意味するɸasita(はした)にnasi(なし)がくっついて強調されたのがɸasitanasi(はしたなし)です。このnasi(なし)は珍しいですが、奈良時代からありました。中途半端であることから「なっとらん(成っていない)」となり、「なっとらん(成っていない)」が強調されたのが、ɸasitanasi(はしたなし)です。無作法であるという意味です。
「水」を意味していたpataのような語も、ずいぶん遠くまで来たものです。
※おそらく、ɸasi(箸)も今回の話に無関係ではないでしょう。「間」を意味したɸasama(はさま)からɸasami(はさみ)が生まれたことを考えると、ɸasi(箸)の背後にもɸasi(間)があると思われます。同じɸasi(間)から生まれたɸasi(橋)とɸasi(箸)がなぜ異なるアクセントを持つようになったのかということですが、ɸasi(橋)とɸasi(箸)は全く別の物であり、経緯が少し違うのでしょう。ɸasi(橋)とɸasi(箸)が同じ場所で同時に生まれたとは考えづらいです。筆者が生まれ育った関東では、「橋」は「し」にアクセントを置き、「箸」は「は」にアクセントを置くのが普通ですが、関西では、「橋」の「は」にアクセントを置き、「箸」の「し」にアクセントを置く逆のパターンを耳にします。
(続く)
参考文献
大野晋ほか、「岩波 古語辞典 補訂版」、岩波書店、1990年。
上代語辞典編修委員会、「時代別国語大辞典 上代編」、三省堂、1967年。
「天(あま)」の語源は「雨(あま)」の語源よりはるかに難しかった、ミャオ・ヤオ語族と仲良しの日本語
筆者にとって、ama*(雨)とama*(天)はずっと気になる存在でした。この二語は、関係がありそうでもあり、関係がなさそうでもあります。
これまでの記事で示してきたように、「水」→「雨」→「落下、下方向、下」という意味変化の超頻出パターンがあるので、ama*(雨)は理解しやすいです。それに比べて理解しづらいのが、ama*(天)です。
結論から言うと、ama*(天)の語源は大変意外です。*ama(天)は、ama*(雨)とは全然違う歴史を持っています。ama*(天)の意外な語源をこれからお話ししますが、その前にミャオ・ヤオ語族の語彙をもっとよく見ておきましょう。
ミャオ・ヤオ語族で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語が日本語に入り、ama*(雨)、umi(海)、una(海)などになったのでした。
当然、奈良時代の日本語のamu(浴む)も、ここに属します。

奈良時代には、abu(浴ぶ)ではなく、amu(浴む)という形が一般的でした。よくあるmとbの間の発音変化です。
bとpまたはɸの間の発音変化も起きやすいです。奈良時代にはaburu(溢る)という形が一般的でしたが、のちにaɸuru(溢る)という形が一般的になりました(現代の日本語には、abureruとahureruという形で両方残っています)。水が荒れ狂ったり、外に出てしまうところから来ているのが、amaru(余る)であり、abaru(暴る)であり、aburu(溢る)です。
唇のところで作る音として、m、b、p、ɸのほかに、wも忘れてはなりません。奈良時代の日本語のawa(泡)、awi(藍)、awo(青)も、明らかに水関連です。
ミャオ・ヤオ語族で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語は、am-という形だけでなく、ab-、ap-、aɸ-、aw-という形でも日本語に残ったということです。溺れていることを表すappuappu(あっぷあっぷ)の語源も、間違いなくここです。
以下の音の間で発音変化が起きやすいことは、頭に入れておかなければなりません。

(ヨーロッパの言語だったら、fとvも考えなければならないところですが、東アジアでは、fとvは一般的ではありません。)
しかし、明らかに水と関係がある上記の語彙に比べると、ama*(天)はなんとも微妙です。ちなみに、奈良時代の日本語には、aɸugu(仰ぐ)という語もありました。aɸugu(仰ぐ)は、もともと上を見ることを意味し、のちに尊敬すること、さらに目上の者になにかを請うこと・求めることを意味するようになりました。aɸugu(仰ぐ)のaɸuは、aɸumuke(仰向け)にも組み込まれているように、「上」を意味し、その古形は日本語の発音の定説によりapu*と推定されます。
ama*(天)とapu*(上)は、形的には上のama*(雨)、appuappu(あっぷあっぷ)、aburu(溢る)、aɸuru(溢る)などとぴったり合いますが、意味的になかなか「水」に結びつきません。
ここから意外な展開に・・・
「水」→「雨」→「落下、下方向、下」という意味変化の超頻出パターンがあるので、「下」を意味する語は簡単に理解できますが、「上」を意味する語はどうしたものかと、筆者もずいぶん悩みました。
筆者の頭に、ある考えがひらめいたのは、uɸe(上)の語源についてあれこれ考えている時でした。ヒントをくれたのは、本ブログでよく引用している三省堂時代別国語大辞典上代編でした。時代別国語大辞典上代編は、奈良時代の日本語のuɸe(上)とkami(上)を比較して、以下のように述べています(上代語辞典編修委員会1967)。
シタに対応するウヘという語が、表面・人の目に触れる所をさすのに対して、カミ・シモは一つづきのものの上下の位置をあらわし、土地の高い所、川の上流、ある地域で中央に近い所、あるいは人間関係における長上を示す。
三省堂時代別国語大辞典上代編には、本当にお世話になっており、感謝しかありません。時代別国語大辞典上代編が指摘しているように、奈良時代の日本人は、uɸeを「上」と書くだけでなく、「表」とも書いていました。
「水」→「雨」→「落下、下方向、下」という意味変化の超頻出パターンがありますが、ここからさらに、「下」→「穴」という意味変化のパターンがあったことを思い出してください。
穴があると、以下の図のようになります。

言われてみると、どうってことないのですが、穴があると、青い部分と赤い部分に「上下関係」が発生するのです。

つまり、今まで「陸地」を意味していた語に、「上」という意味が生じるのです。ミャオ・ヤオ語族で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語は、「雨」を意味するようになったが、その一方で、「陸地」も意味していたということです。筆者がいつも描いているあの図です。

水を意味していた語が、その横の部分を意味するようになるパターンです。「水」を意味していたamaが、「陸地」を意味するようになり、上で説明した過程を経て、「上」を意味するようになるのです(水を意味していたapuも同様です)。これが、日本語のama*(天)の語源です。
前回の記事で、無関係な二語としてkawa(川)とkawa(皮)を挙げましたが、実は、kawa(川)とkawa(皮)には関係があります。
「水」を意味していたkapaのような語は、「川」を意味するようになったが、その一方で、「陸地」も意味していたのです。このkapaのような語が、上で説明した過程を経て、「表面」を意味するようになったのです。これが、日本語のkaɸa(皮)の語源です。
奈良時代の日本人は、体の表面部分のことを、kaɸa(皮)と言うこともあれば、kaɸabe(皮)と言うこともありました。このkaɸa(皮)とkaɸabe(皮)は、kaɸa(川)とkaɸabe(川辺)と同源なのです。
奈良時代の日本人がなぜuɸeを「上」と書いたり「表」と書いたりしていたのか、理解できたでしょうか。uɸa*(上)もuɸabe(上辺)も、元を辿れば、ミャオ・ヤオ語族で「水」をしたam、ab、ap、aw、um、ub、up、uw、om、ob、op、owのような語から来ているのです。
kaɸa(皮)の語源が上の通りなら、ɸada(肌)の語源も俄然怪しくなってきます。
batyabatya(バチャバチャ)、basyabasya(バシャバシャ)、ɸata(辺、端)、ɸata(畑)などの語から、「水」を意味していたpataのような語が、「陸地」を意味するようになったことがはっきりと窺えるからです(日本語に近縁な言語が、「水」のことをmat-、mit-、mut-、met-、mot-と言ったり、bat-、bit-、but-、bet-、bot-と言ったり、pat-、pit-、put-、pet-、pot-と言ったりしていたということです)。

「水」を意味していたpataのような語が、「陸地」を意味するようになり、上で説明した過程を経て、「表面」を意味するようになったと考えられます。これが、日本語のɸada(肌)の語源です。
ここで、ちょっと気になることがあります。先ほどɸata(畑)という語を挙げましたが、ɸatake(畑)という形もあります。ɸatake(畑)のkeは乙類なので、ɸataka*(畑)という古形が推定されます。
このɸata(畑)とɸataka*(畑)は、いわくありげです。なぜなら、ɸada(肌)とɸadaka(裸)という語があるからです。この関係は、先ほどのkaɸa(川)とkaɸabe(川辺)、kaɸa(皮)とkaɸabe(皮)の関係を思い起こさせます。
以下のようなことがあったのではないかと思われます。
まず、「水」を意味するpataのような語があります。そして、この語が横の部分を意味するようになります。
その一方で、「水」を意味するkapaのような語があります。そして、この語が横の部分を意味するようになります。

ɸata(辺、端)とkaɸa(側)という語が実在するわけですから、上の変化は実際にあったと考えられる変化です。
ɸata(辺、端)のほかに異形と見られるɸeta(辺、端)という語もあり、このɸata(辺、端)とɸeta(辺、端)は、特に複合語でɸaとɸeに短縮されていました。ここから来ているのが、kaɸaɸe(川辺)やumiɸe(海辺)です(のちにkaɸabe(川辺)とumibe(海辺)が一般的になります)。
上のような短縮はよく起きており、midu(水)とumi(海)も、特に複合語でmiとuとして現われることがありました。おそらく、kaɸa(川)(あるいはkapa*(川))も、特に複合語でkaとして現われることがあったのではないかと思われます。

なにが言いたいかというと、左下の「水」を意味したkapaのような語と、右上の「横の部分」を意味したpataのような語が組み合わさって、短縮したのが、kaɸabe(皮)であり、左上の「水」を意味したpataのような語と、右下の「横の部分」を意味したkapaのような語が組み合わさって、短縮したのが、ɸadaka(裸)ではないかということです。ɸataka*(畑)も、ɸadaka(裸)と同様です。
「下」を意味する語の語源に比べると、「上」を意味する語の語源は、なんというか、トリッキーですね。筆者も驚きました。
ポイントは、「水」を意味していた語が、「陸地」を意味するようになり、上で説明した過程を経て、「上」または「表面」を意味するようになるということです。
※今回の記事では、ama、pata、kapaが水の横の部分を意味するようになる場面が出てきました。

上の語はどことなく、amu(虻)(のちにabuに変化)、ɸati(蜂)、ka(蚊)を思わせます。おそらく、偶然ではないでしょう。
真ん中のpataを見てください。「横」を意味しています。そして、日本語にpatapata(パタパタ)、batabata(バタバタ)、basabasa(バサバサ)という語があります。「横」を意味していた語が、「羽、翼、飛ぶ」の意味領域に進出するのだろうなと察しがつきます。ɸato(鳩)の語源もここでしょう。
上の構図に当てはまらないのは、ɸaɸe(蝿)です。ɸaɸe(蝿)のɸeは乙類なので、ɸaɸa*という古形が推定されます。動詞のɸaɸu(這ふ)から来たのでしょう。

蝿は、確かに飛びますが、食べ物の表面に付着して這いまわるイメージが強いのでしょう。ちなみに、英語のfly(飛ぶ)とfly(蝿)は同源です。
補説1
過去の記事の修正、aɸu(合ふ)とaɸu(会ふ)
日本語のaɸu(合ふ)とaɸu(会ふ)については、このブログを書き始めた頃に、全く見当違いの説明をしてしまったので、ここで修正させてください。
奈良時代の日本語のaɸu(合ふ)は、四段活用です。

未然形はaɸaで、このaɸaがなにを意味していたのか考えなければなりません。他の語と同様に、apa*という古形が推定されます。
ミャオ・ヤオ語族で「水」を意味したam、am、an、aŋ、um、un、uŋ、om、on、oŋのような語が、am-という形だけでなく、ab-、ap-、aɸ-、aw-という形でも日本語に入ったという話をしました。日本語の語彙を見る限り、以下のような構図があったと見られます。

「水」を意味していた語が、「横の部分」を意味するようになったところです。本ブログでお話ししてきたように、ここから、「1」を意味することも、「2」を意味することもできるし、「一方」を意味することも、「もう一方」を意味することも、「両方」を意味することもできます。
「2」に近いですが、「対、組、ペア」を意味することもできます。apa*は「対、組、ペア」を意味していて、それから作られた動詞のapu*は「対になること、組になること、ペアになること」、あるいは場合によっては、「一体になること」を意味していたと見られます。抽象的な図ですが、なにかが二つあって、それが以下のようになることを意味していたと見られます。

例えば、磁石のN極とS極だったら、こうなりますが、N極とN極だったら、こうはなりません。前者の場合には、「aɸu(合ふ)」と言い、後者の場合には、「aɸanu(合はぬ)」と言うわけです。
補助動詞として使われたaɸu(合ふ)は、動作がいっしょに行われること、動作が双方向に行われることを意味していましたが、これは、apa*が「対」を意味し、aɸuが「対になること」を意味していたことを考えれば、納得できるでしょう。
二者が接近することあるいは接近していることを意味したaɸi(合ひ)と場所を意味するta(konata(こなた)やkanata(かなた)のta)がくっついたのが、aɸida(間)でしょう。
奈良時代の日本語には、aɸu(合ふ)のほかに、aɸu(敢ふ)という動詞もありました。現代でも、多少無理をする時に「敢えて」と言いますね。aɸu(敢ふ)は、下二段活用です。

aɸu(敢ふ)は、なにかに対抗すること・抵抗することを意味していました。この動詞も、上のapaの図から来たと考えられます。
apaの図のところで、「1」を意味することも、「2」を意味することもできるし、「一方」を意味することも、「もう一方」を意味することも、「両方」を意味することもできると述べました。この中の「もう一方」から、「反対」や「逆」のような意味が生まれてくるのです。だから、aɸu(敢ふ)という動詞は、対抗・抵抗という意味を持っていたのです。
例えば、saka(逆)もこのパターンです。

saka*(酒)から、「水」を意味したsakaのような語があったことが窺えます。この語が「横の部分」を意味するようになります(水と陸の境界を意味するsaka(境)にもなりました)。ここから、「一方」を意味することも、「もう一方」を意味することも、「両方」を意味することもできますが、実際には、「もう一方」を意味するようになり、「反対」や「逆」のような意味が生まれてきます。これが、saka(逆)の語源です。反対側、反対方向、逆側、逆方向を意味します。sakaɸu(逆ふ)とsakaru(逆る)という動詞も作られましたが、これらは廃れてしまいました。
「水」を意味する語から「反対」や「逆」のような意味が生まれてくるのも重要なパターンなので、覚えておいてください。
補説2
古い頭、kabu(頭)とkaube(頭)
今回の記事のkaɸa(川)とkaɸabe(川辺)、kaɸa(皮)とkaɸabe(皮)に関連して、もう一つ気になることがあります。
それは、kabu(頭)とkaube(頭)です。
前に、「頭(あたま)」の語源、仇(あだ)の意味に関する考察からという記事を書きましたが、atama(頭)という語が現れるのは、室町時代からで、しかも最初は、頭というより、赤ん坊の頭の前のほうに見られるへこみを意味していました。室町時代より前に頭を意味していた語はいくつかありますが、その中にkabu(頭)とkaube(頭)がありました。
kabu(頭)は奈良時代からあり、kaube(頭)は平安時代から現れます。平安時代は、語中のɸがwに変化した時期です。ただ、この変化によって、kaɸaはkawaになることができますが、kaɸuはkawuになることができません。日本語にwuという音はないからです(末尾の注も参照)。kaɸuはkauにならざるをえません。
現代の日本人がgabunomi(がぶ飲み)、gabugabu(がぶがぶ)と言っていることから、「水」を意味するkapu*という語があったと考えられます。「水」を意味するkapa*の異形でしょう。
kapa*のほうは、kaɸaとkaɸabeになり、「陸地」を意味するようになりました。kapu*のほうも、kaɸu*とkaɸube*になり、「陸地」を意味するようになったら、どうでしょうか。「陸地」を意味していた語が「表面」または「上」を意味するようになるのは、頻出パターンです。
kaɸaとkaɸabeは、「表面」を意味するようになり、さらに「皮」を意味するようになりましたが、kaɸu*とkaɸube*は、「上」を意味するようになり、さらに「頭」を意味するようになったのではないかと思われます。
「下」を意味していた語が「足」を意味するようになり、「横」を意味していた語が「手、腕」を意味するようになるのが人類の言語の超頻出パターンなら、「上」を意味していた語が「頭」を意味するようになるのは自然です(「真ん中」を意味していた語はonaka(お腹)になっています)。
kaɸu*とkaɸube*の場合は、発音の変化がちょっと複雑で、kaɸu*が濁ってkabu(頭)になった時に、kaɸube*は濁ってkabube×にならなかったと見られます。kaɸube*のまま残ると、平安時代の変化でkawube×になることができず、kaube(頭)になります。
※奈良時代の時点で、wa、wi、we、woはありましたが、wuはありませんでした(一般に、人類の言語において、wという子音は消滅しやすいです)。奈良時代の日本語には、uu(植う)とuu(飢う)という語がありました。

昔の日本語は母音の連続を許さなかったので、uu(植う)とuu(飢う)は異例です。おそらく、uwu*という古形があったでしょう。
ミャオ・ヤオ語族で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語が、am-、um-、om-という形だけでなく、ab-、ap-、aw-、ub-、up-、uw-、ob-、op-、ow-という形でも日本語に入ったことをお話ししました。uu(植う)とuu(飢う)の未然形のuweも、そこから来たと見られます。「水」→「雨」→「落下、下方向、下」の超頻出パターンです。
uweは「下」を意味していて、そこから「地下・地中」を意味するようになってできたのがuu(植う)(古形uwu*)で、「衰弱すること、死ぬこと」を意味するようになってできたのがuu(飢う)(古形uwu*)でしょう。英語のstarveは、「死ぬこと」を意味していましたが、意味が特殊化し、「餓死すること、飢えること」を意味するようになりました。日本語のuu(飢う)も、このパターンと考えられます。
補説3
kabu(頭)と関係がありそうなkabuto(かぶと)
kabu(頭)と無関係とは思えない語として、kabuto(かぶと)があります(kabuto(かぶと)のtoは、甲類と乙類の間で揺れていました(上代語辞典編修委員会1967)。筆者には、このことも非常に重要に思われます)。kabu(頭)とkabuto(かぶと)は、無関係とは思えないですが、どういう関係があるのかは難問です。
しかし、今回の記事で見たkaɸa(皮)とkaɸabe(皮)、ɸada(肌)とɸadaka(裸)などのケースが参考になると思います。
上で説明したように、kabu(頭)は、gabunomi(かぶ飲み)/gabugabu(かぶがぶ)から、もともと「水」を意味していたと推測されます。問題は、kabutoのkabuの部分ではなく、toの部分です。以下のようになっていた可能性が高いです。

※上の図のkabuは、kaɸu*であったかもしれません。いずれにせよ、kaɸa(川)と同源です。
kabuは「水」を意味し、toは「横の部分」を意味していただろうということです。
minato(港)の語源は、「水」を意味するmiと、noと同じ働きをする助詞のnaと、「門」を意味するtoと説明されてきましたが、toを「門」と決めつけるのは問題です。
奈良時代には、to(門)という語だけでなく、場所を意味するto(処)という語もありました。
minato(港)は、「水」を意味するmiと「横の部分」を意味するtoがくっついたもので、水に隣接する場所と解釈したほうが、minato(港)以外の語彙も理解しやすいのです。
例えば、yamato(大和)です。前方後円墳とは何だったのか、その始まりも重要だが、その終わりも重要の記事で、巨大前方後円墳が続々と作られ始めた場所を見ました。同記事の地図を見れば一目瞭然ですが、日本という国の発祥の地は、もろに三輪山の麓にあります。yamato(大和)は、「山」を意味するyamaと「横の部分」を意味するtoがくっついたもので、「山際」だったのだとわかります。
minato(港)のtoは甲類で、yamato(大和)のtoは乙類です。これも非常に重要です。kabuto(かぶと)のtoも、甲類と乙類の間で揺れていたからです。「(水の)横の部分」を意味するtoという語があったが、このtoは甲類と乙類の間で揺れていたということです。
「水」を意味するkabuと「横の部分」を意味するtoがくっついてkabutoができ、「陸地」を意味していたが、今回の記事で説明した過程を経て、「上」を意味するようになり、さらに「頭」を意味するようになったのです。
kabu(頭)とkabuto(かぶと)は、今回の記事で見たkaɸa(皮)とkaɸabe(皮)、ɸada(肌)とɸadaka(裸)などと同様の歴史を持っているということです。
※「横の部分」を意味したtoも、さらにその前は、「水」を意味していたにちがいありません。

日本語のtokoro(所)は、「水」を意味したtoと「横の部分」を意味したkoroがくっついたものでしょう。tokoro(所)は、陸地、土地、場所を意味していたのです。
日本語で、「6時」ではなく、「6時頃」と言ったら、どう意味が変わるでしょうか。「6時の近く」を意味するのではないでしょうか。koro(頃)もまた、水の近くを意味していたのです。
参考文献
上代語辞典編修委員会、「時代別国語大辞典 上代編」、三省堂、1967年。
「生意気(なまいき)」とは何か、誰もが違和感を覚える「舐める」と「ナメる」
前回の記事でタイ系言語の話をたくさんしたので、関連する話題としてnamaiki(生意気)とnameru(ナメる)を取り上げましょう。後者は、「ナメんなよ」のnameru(ナメる)です。日本語を母語とする人でも、「この語はなんだろう」と思ったことがあるのではないでしょうか。
人間の言語の語彙が形成されていく過程は必ずしも単純ではなく、ちょっと入り組んだ現象が起きることもあります。
日本語にkawa(川)という語とkawa(皮)という語があります。皆さんは、この二語を見てどう思うでしょうか。「発音が同じなだけで、関係ないな」と思うでしょう。kawa(川)とkawa(皮)の場合はそれで終わるのですが、それで終わらない場合もあるのです。
ある意味(意味A)を持つnamaという語があったとしましょう。そして、別の意味(意味B)を持つnamaという語があったとしましょう。

最初は、左のnamaと右のnamaは別物として存在しています。しかし、時間が経つうちに、Aという意味がBという意味に干渉し始める、あるいは、Bという意味がAという意味に干渉し始めることがあります。
Aという意味でnamaという語を用いていたが、そこにBという意味がうっすらと漂い始める、あるいは、Bという意味でnamaという語を用いていたが、そこにAという意味がうっすらと漂い始めるということです。
この現象は、人間の言語を面白くしますが、難しくもします。
namaという語になにが起きたのか、歴史を振り返ってみましょう。
●nameru(ナメる)
タイ系言語で「水」を意味したnam-、nim-、num-、nem-、nom-のような語が、「下」を意味するようになったことをお話ししました(「水」→「雨」→「落下、下方向、下」の変化です)。namaあるいはnameのような語が「下」を意味していたわけです。
nameru(ナメる)は、日本語のスラング(俗語)ですが、「侮る、見下す、見くびる」の類です。「侮る、見下す、見くびる」のような語を見ればわかるように、nameru(ナメる)は「下に見ること」を意味していたと考えられます。namaあるいはnameのような語が「下」を意味していたところから生まれてきた語です。最初は、純粋に下を見ることを意味し、侮蔑の意味はなかったのかもしれません。奈良時代の時点ですでに、namesiという形容詞が存在し、無礼な態度を意味していました。
※anadoru(侮る)は、奈良時代にはanaduruという形がよく使われ、anaturu、anadoru、anatoruという形が少し見られましたが、いずれにせよ、意味を考えると、anadoru(侮る)のanaの部分は「穴」というより「下」を意味し、doruの部分は「取ること(捉えること)」を意味していたと見られます。
mikudasu(見下す)のkudaは、kudaru(下る)とkudasu(下す)という語があるので、「下」を意味していたことは明らかです。mikubiru(見くびる)のkubiは、なんでしょうか。このkubiも、「下」を意味していたようです。mikubiru(見くびる)は、miorosu(見下ろす)、misageru(見下げる)、mikudasu(見下す)に並べられる語だということです。「下」を意味していたkubiが「首」を意味するようになる過程は、補説を参照してください。
namaあるいはnameのような語が「下」を意味していたことさえわかれば、「下に見ること」を意味するnameruは、無理なく理解できるでしょう。
もっと微妙なのが、namaiki(生意気)です。
●namaiki(生意気)
すでに述べたように、タイ系言語で「水」を意味したnam-、nim-、num-、nem-、nom-のような語は、「水」→「雨」→「落下、下方向、下」と意味変化し、「下」を意味するようになりました。
しかし、その一方で、「湿っていること」も意味するようになりました。

ここから特に、魚や肉が、まだ焼いたり干したりしておらず、水っぽい状態であることを意味するようになりました。そこからさらに、処理などの過程を経ていない状態であること、途中の状態であること、完成していない状態であることを意味するようになりました。
namaiki(生意気)のnamaには、「下」という意味も含まれていますが、単純にそれだけではなく、「過程を経ていない、途中である、完成していない」という意味も含まれているのです。だから、子どもや未熟者が意気がった時によく、namaiki(生意気)という言葉が飛んでくるのです(現代の日本語にnamamono(生物)という語がありますが、かつてはnamamono(生者)という語もあり、一人前でない者を意味していました)。
単純に相手を見下すだけなら、namaiki(生意気)以外にも表現はたくさんあるでしょう。namaiki(生意気)が独特の意味、ニュアンス、語感を持っているのは、上のような歴史があるからなのです。
kubi(首)の語源
mikubiru(見くびる)に組み込まれているkubiは、「下」を意味していた語のようだと述べました。miorosu(見下ろす)、misageru(見下げる)、mikudasu(見下す)と比較すれば、妥当な推論でしょう。
「下」を意味していたkubiが「首」を意味するようになる過程は難しくありません。kubo(窪)やkubire(くびれ)のような語を見ればわかります。「下」を意味していた語が「穴、くぼみ、へこみ」を意味するようになったのです。
奈良時代の日本語には、kubi(首)の同義語として、una(頸)という語もありました。岩波古語辞典は、首を意味するuna(頸)とうしろを意味するsiri(後)がくっついて短縮したのがunazi(うなじ)であると考えていますが、これはその通りでしょう(大野1990)。
kubi(首)が「下」を意味していたのと同様に、una(頸)も「下」を意味していたと考えられます。
ミャオ・ヤオ語族で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語が、「水」→「雨」→「下」と意味変化し、ana*もuna*も「下」を意味していたのでしょう(anadoru(侮る)の話を思い出してください)。そこから、ana*は「穴、くぼみ、へこみ」に落ち着き、una*は(「穴、くぼみ、へこみ」を経て)「首」に落ち着いたのでしょう。
他の身体部位もそうですが、kubi(首)もuna(頸)も、もともと身体部位を意味していた語ではないということです。
タイ系言語とミャオ・ヤオ系言語が物語る日本語の歴史の核心部分
タイ系言語とミャオ・ヤオ系言語から日本語に大量の語彙が入ったことをお話ししていますが、その中で、日本語のnami(波)とumi(海)は、特に注目に値します。
タイ系言語で「水」を意味したnam、nim、num、nem、nomのような語から来たのが、日本語のnami(波)です。
ミャオ・ヤオ系言語で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語から来たのが、日本語のumi(海)です。
タイ系言語で「水」を意味したnam、nim、num、nem、nomのような語が、日本語に入ろうとしたが、日本語にはmidu(水)という語があるので、「波」を意味するようになったのです。
ミャオ・ヤオ系言語で「水」を意味したam、an、aŋ、um、un、uŋ、om、on、oŋのような語が、日本語に入ろうとしたが、日本語にはmidu(水)という語があるので、「海」を意味するようになったのです。
よくよく考えると、これは非常に重要なことです。
「水」を意味することができなくなったから、「波」を意味しよう、これが可能なのは、波がある場所です。波がない場所で、「波」を意味する語にはなれないでしょう。
「水」を意味することができなくなったから、「海」を意味しよう、これが可能なのは、海がある場所です。海がない場所で、「海」を意味する語にはなれないでしょう。
日本語の歴史の核心的な部分になりますが、海があり、波がある場所で、タイ系言語とミャオ・ヤオ系言語から直接、日本語に語彙が入ったということです。
「水」を意味していた語が「口」を意味するようになることは、可能でしょうか。可能です。「水」→「雨」→「下」という意味変化は超頻出パターンですが、「下」→「穴」→「口」という意味変化も超頻出パターンです(「下」→「穴」→「口」という意味変化については、「口(くち)」の語源の記事で詳しく論じました)。すなわち、「水」→「雨」→「落下、下方向、下」→「穴」→「口」という黄金パターンの完成です。

しかし、「水」を意味していた語がいきなり「口」を意味するようになることはできません。
例えば、日本語のnamu(舐む)(未然形name)とumeku(呻く)という語について考えてみましょう。
nameという形からして、タイ系言語で「水」を意味した語が、「水」→「雨」→「落下、下方向、下」→「穴」→「口」と意味変化し、この「口」を意味した語から生まれたのが、namu(舐む)であると考えられます(正確には、最後に「口」→「舌」という意味変化があったと考えられます。「口」を意味するnamaあるいはnameのような語があったことは、「名前(なまえ)」とは何か、平仮名と片仮名についてもう一言の記事でお話ししたna*(音)からも推測できます。na*(音)がnaru(鳴る)/nasu(鳴す)、naku(鳴く)、na(名)、ne(音)などになったのでした。このna*(音)も、「口」を意味していた語でしょう。namaru(訛る)/namari(訛り)も、「口」と「音」に関係があるのかもしれません。ひょっとしたら、先ほど見たような「意味の干渉」が起きたのかもしれません。これは本当に、興味深い現象なのです。

namaru(訛る)/namari(訛り)は、「話すこと、音」を意味していますが、それが普通でないところに特徴があります。左のnamaは「下」を意味し、そこからnamaru(鈍る)などの語が生まれました。左の「下」から「調子が落ちる、調子が崩れる、調子が悪くなる」のような意味が生じると、右の「音」に干渉しそうでもあります。日本語では、namという形が許されないので、nama、nameもしくはnaのような形になります。「口」→「舌」という意味変化は、下(した)と舌(した)、そこには奇妙で怪しい関係が・・・の記事でも取り上げました)。
umeという形からして、ミャオ・ヤオ系言語で「水」を意味した語が、「水」→「雨」→「落下、下方向、下」→「穴」→「口」と意味変化し、この「口」を意味した語から生まれたのが、umeku(呻く)であると考えられます。
※「下」を意味するnamaのような語と、「下」を意味するumaのような語があったことは、前回の記事で確認済みです。
しかし、「水」からnamu(舐む)に至る道のりと、「水」からumeku(呻く)に至る道のりは、とても長いです。
namu(舐む)は、究極的根源まで遡れば、タイ系言語の「水」から来ているようですが、namu(舐む)という語が、タイ系言語から日本語に直接入ったかどうかは、わからないのです。間に他の言語を挟んだ可能性が十分にあります。
umeku(呻く)は、究極的根源まで遡れば、ミャオ・ヤオ系言語の「水」から来ているようですが、umeku(呻く)という語が、ミャオ・ヤオ系言語から日本語に直接入ったかどうかも、わからないのです。間に他の言語を挟んだ可能性が十分にあります。
しかし、nami(波)とumi(海)は、namu(舐む)とumeku(呻く)とは決定的に違います。
「水」を意味していた語が、いくつもの意味変化を経て、「波」を意味するようになった、あるいは、「水」を意味していた語が、いくつもの意味変化を経て、「海」を意味するようになった、そういう間接的なストーリーは考えられないのです。「水」から「波」への意味変化も、「水」から「海」への意味変化も、ダイレクトなのです。
先ほど強調したように、海があり、波がある場所で、タイ系言語とミャオ・ヤオ系言語から直接、日本語に語彙が入ったのです。
タイ系民族とミャオ・ヤオ系民族を含め、中国南部から東南アジア大陸部にかけて分布している民族と日本人の文化的な共通性・類似性は、これまでにも指摘されてきました(鳥越1992)。文化的な共通性・類似性の指摘自体は、極めて適切です。しかし、そこから、日本語は中国南部から東南アジア大陸部にかけての地域から来たと結論するのは、適切ではありません。今、日本人が遠くに見ているタイ系言語、ミャオ・ヤオ系言語、ベトナム系言語が、中国東海岸地域に(も)存在したのです。日本語があっちにあったのではなく、タイ系言語、ミャオ・ヤオ系言語、ベトナム系言語がこっちに(も)あったのです。
東アジアの運命を決定した三つ巴、二里頭文化と下七垣文化と岳石文化の記事では、内陸に存在した巨大な殷が中国東海岸地域に侵攻してきたことをお話ししました。激動の時代、うまくいかなくなったアワとキビの栽培、うまくいかなくなったイネの栽培の記事では、気候変動のために朝鮮半島で稲作がうまくいかなくなり、朝鮮半島の人口が激減したことをお話ししました。どちらも、そこに住んでいた人々を広く襲った大事件であり、日本語とその近縁言語の話者だけが、中国東海岸地域から朝鮮半島に移動し、朝鮮半島から日本列島に移動したとは全然限りません。事件の規模を考えれば、タイ系言語、ミャオ・ヤオ系言語、ベトナム系言語の話者(の一部)が、中国東海岸地域から朝鮮半島に移動した可能性、さらに朝鮮半島から日本列島に移動した可能性すらあるのです。
参考文献
大野晋ほか、「岩波 古語辞典 補訂版」、岩波書店、1990年。
鳥越憲三郎、「古代朝鮮と倭族 神話解読と現地調査」、中央公論新社、1992年。