
現代人のDNAだけでなく、古代人のDNAまで調べられるようになったのは、頼もしい限りです。東南アジアは高温多湿で、古代人の遺骨の保存状態がよくない点は不利ですが、それでも、驚くべき研究が次々に発表されています。
まずは、M. Lipson氏らの図をどーんと掲げましょう(図はLipson 2014より引用)。

東南アジアの歴史に関しては、研究者の間で意見が一致していることと、意見が分かれていることがあります。Lipson氏らの図は、現時点で研究者の間で意見が一致していることを要領よくまとめたものです。
矢印が多いので、順に説明しましょう。赤、オレンジ、青、緑の矢印は、人の動きを示しています。赤とオレンジの矢印は非常に古い矢印で、青と緑の矢印は比較的新しい矢印です(矢印のルートは完全に正確ではありません)。そして、各棒は、現代の各地の民族のDNAを示しています。
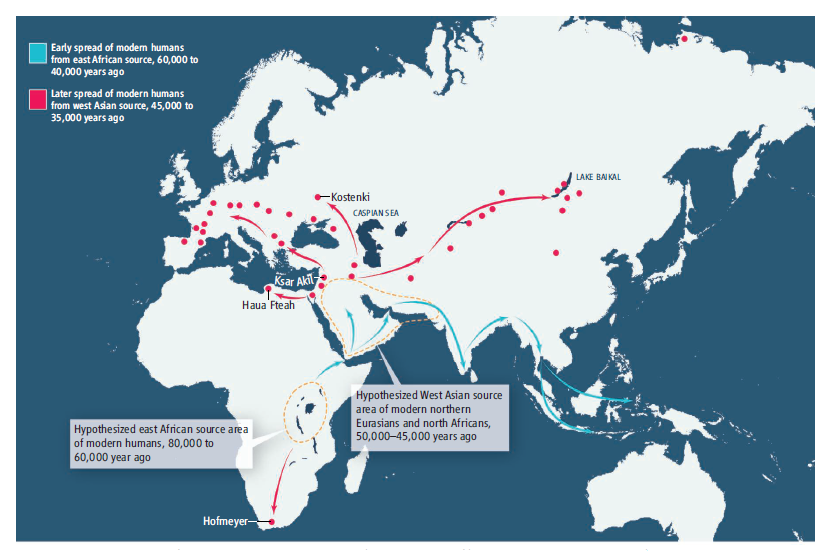
赤とオレンジの矢印は、人類がアフリカを出て、中東と南アジアを通過し、東南アジアに到達した頃のものです。つまり、5万年前頃のものです。
当時は現在と地形が全然違うので、もう一度確認しておきましょう(図はWikipediaより引用)。
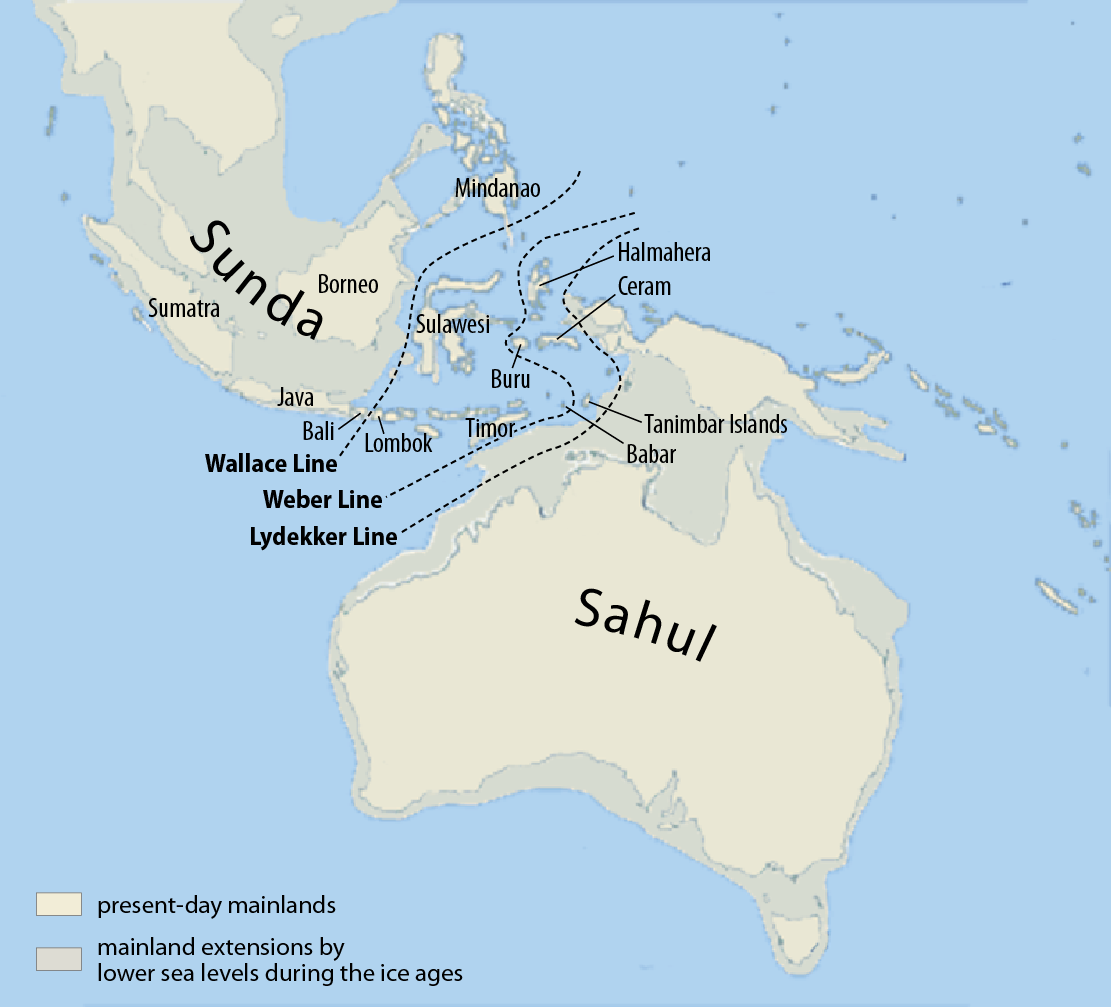
スンダランドとサフルランドという巨大な陸が相対していたことは、以前にお話ししました。
インドネシアには、スマトラ島、ジャワ島、ボルネオ島(カリマンタン島ともいいます)、スラウェシ島という四つの大きな島がありますが、スマトラ島、ジャワ島、ボルネオ島は、大陸とつながっていました。スラウェシ島は、大陸とつながっていませんでした。
フィリピンは、大陸とつながっていそうで、つながっていませんでした。
フィリピンとインドネシアのスラウェシ島が大陸につながっていなかったというのは重要なポイントなので、覚えておいてください。
Lipson氏らの図に戻りましょう。
赤とオレンジの矢印は、人類がアフリカから東南アジアに到達した頃のものですが、赤の矢印は、スンダランドからフィリピンに向かう動き、オレンジの矢印は、スンダランドからサフルランド(パプアニューギニアとオーストラリア)に向かう動きを示しています。スンダランドから先は、海を渡らなければなりませんでした。
フィリピンには、背が低く、色が黒く、髪が縮れているいくつかの少数民族が住んでおり、これらの少数民族はまとめて、「ネグリト」と呼ばれています(画像はPreda Foundation様のウェブサイトより引用)。


前回の記事では、オーストロネシア語族の言語を話す人々が、5000年前頃から、中国南東部を離れて、台湾、フィリピン、インドネシア、マレーシアに進出していったことをお話ししました。Lipson氏らの図の青の矢印が、オーストロネシア語族の言語を話す人々の動きです。
オーストロネシア語族の言語を話す人々がやって来る前からフィリピンにいたのが、ネグリトです。「ネグリト」という名は、スペイン人が付けたもので、スペイン語で「小さな黒人」を意味するnegritoから来ています(フィリピンがスペインの植民地になった経緯については、補説を参照)。
ネグリトは、長い間、人類学者、生物学者、考古学者にとって大きな謎でしたが、人間の全DNAが容易に調べられるようになって、ようやくその素性が明らかになってきました。
Lipson氏らの図のフィリピンのところを見てください。Ayta(またはAeta)というのは、フィリピンで最もよく知られている少数民族です。Tagalogというのは、現在のフィリピンで多数派になっている普通のフィリピン人です。AytaのDNAを見ると、赤の矢印の人々のDNAの割合が高いですが、青の矢印の人々のDNAもある程度入っています。TagalogのDNAを見ると、青の矢印の人々のDNAの割合が高いですが、赤の矢印の人々のDNAもある程度入っています。中国南東部からやって来た人々とフィリピンの原住民は、ある程度混ざり合ったということです。
ネグリトに関しては、もう一つ重要なことがわかりました。
以前に、デニソワ人の話をしたことがありました。デニソワ人は、ネアンデルタール人とは別種の古人類です。デニソワ人は2008年に発見されたばかりなので、知らない人も多いかと思います。詳しくは、本ブログの過去の記事を参照してください。
ネアンデルタール人が現生人類と交わったのと同様に、デニソワ人も現生人類と交わったわけですが、特にパプアニューギニアとオーストラリア(アボリジニ)の人々のDNAに、デニソワ人のDNAが顕著に入っていることが明らかになりました。このように、まずはパプア人とアボリジニのDNAに注目が集まりましたが、すぐにネグリトのDNAも注目されました。パプア人、アボリジニ、そしてネグリトを含む東南アジアの人々のDNAに、デニソワ人のDNAがどのくらい入っているか見てみましょう(図はTeixeira 2021より引用)。

パプア人とアボリジニに入っているデニソワ人のDNAの量は同じくらいで、その量を100%としています。そして、東南アジアの人々にデニソワ人のDNAがどのくらい入っているか見ています。
見ての通り、かつてのサフルランドの地域にいる人々には、デニソワ人のDNAが顕著に入っていますが、かつてのスンダランドの地域にいる人々には、デニソワ人のDNAはほとんど入っていません。そして、かつてのサフルランドとスンダランドの間の地域にいる人々には、デニソワ人のDNAがほどほどに入っています。
これは、どう解釈したらよいのでしょうか。
結論を先に言ってしまうと、スンダランドとサフルランドの間の地域には、サフルランドの人々と同系統の人々が住んでいたということです。
しかし、スンダランドとサフルランドの間の地域には、のちにオーストロネシア語族の人々とオーストロアジア語族(ベトナム系言語)の人々が流入してきたのです。
さりげなく述べましたが、オーストロネシア語族の人々だけでなく、オーストロアジア語族(ベトナム系言語)の人々も流入してきたのです。
Lipson氏らの図を見ると、青の矢印のほかに、緑の矢印がありますね。青の矢印は、オーストロネシア語族の人々の流入で、緑の矢印は、オーストロアジア語族(ベトナム系言語)の人々の流入です。DNA研究によって、両方の流入があったことが明らかになったのです(インドネシア西部の人々のDNAを見ると、青の矢印の人々のDNAだけでなく、緑の矢印の人々のDNAも多いです。農耕は、中国の長江下流域から台湾に伝わりましたが、その一方で、長江中流・上流域からインドシナ半島(ベトナム、カンボジア、タイ、ラオスなどがあるところ)にも伝わりました)。
オーストロアジア語族(ベトナム系言語)は、東南アジア大陸部の歴史では大いに注目されてきましたが、東南アジア島嶼部の歴史ではほとんど注目されてこなかったので、これは意外な事実です。
先ほどのTeixeira氏らの図から、かつての東南アジアの歴史が窺えます。
デニソワ人のDNAの広がり具合からして、スンダランド東海岸から各島々に渡るあたりで、現生人類(の一部)とデニソワ人の本格的な交わりがあった可能性が高そうです。
Lipson氏らの図で、フィリピンの各民族のDNAを見ます。そして、青の矢印の人々のDNAがどのくらい入っているか見ます。次に、Teixeira氏らの図で、フィリピンの各民族のDNAを見ます。そして、これらの民族に入っているデニソワ人のDNAが、パプア人とアボリジニに入っているデニソワ人のDNAに比べて、どのくらい減っているか見ます。Lipson氏らのデータとTeixeira氏のデータは、見事に一致しています。オーストロネシア語族の人々のDNAが入った分(割合)だけ、デニソワ人のDNAが減っているのです。
フィリピンと同様のことが、インドネシア東部についてもいえます。オーストロネシア語族の人々とオーストロアジア語族(ベトナム系言語)の人々のDNAが入った分(割合)だけ、デニソワ人のDNAが減っているのです。
つまり、オーストロネシア語族の人々とオーストロアジア語族(ベトナム系言語)の人々のDNAが入る前は、デニソワ人のDNAの量はパプア人とアボリジニと同じくらいあったということです。
Lipson氏の図のオレンジの矢印の流れを汲んでいるので、「かつてのインドネシア東部の住民」は、「パプア人」と「アボリジニ」と近い関係にありそうです。
Lipson氏の図の赤の矢印の流れを汲んでいるので、「かつてのフィリピンの住民」は、「かつてのインドネシア東部の住民」、「パプア人」、「アボリジニ」とやや近い関係にありそうです。

前回の記事では、オーストロネシア語族の言語を話す人々は、中国南東部を離れた後、台湾、フィリピン、インドネシア、マレーシアの原住民と出会ったが、その原住民はどんな人たちだったのかと述べました。
「かつてのフィリピンの住民」と「かつてのインドネシア東部の住民」は、パプア人とアボリジニに(やや)近そうだなと推測できます。
「かつての台湾の住民」、「かつてのインドネシア西部の住民」、「かつてのマレーシアの住民」はどんな人たちだったのでしょうか。
最近、この問題を考えるうえで気になる研究が発表されました。(続く)
補説
大航海時代、コロンブス、バスコダガマ、マゼラン
フィリピンのネグリトの話のところで、スペインが出てきました。フィリピンとスペインが結び付かない人がいるかもしれないので、一応書いておきます。
コロンブスの時代のヨーロッパの人たちは、邪魔・妨害をする中東を経由せずに、その向こうの東方の品物を手に入れようとしていました。そこで、陸路の代わりに海路に目が向けられました。しかし当時は、ヨーロッパから海路で東方に行けるかどうか不明でした。
コロンブスは、地球は丸いと信じていたので、ヨーロッパから西に進んでインドに到達できると考えていました。実際に、スペインの援助を受けてインドを目指します。しかし、中米に到着してしまいます。コロンブスは、「自分はインドに到達した」と信じていました。その後、アメリゴベスプッチの探検によって、それがインドではなく新大陸であることが明らかにされました。それでも、コロンブスは、「自分はインドに到達した」と信じていました。
そうこうしているうちに、ポルトガルのバスコダガマが、ヨーロッパからアフリカ大陸の南端を回ってインドに到達してしまいます。こっちは本物のインドです。
ポルトガルに先を越されたスペインは、新しい航路を確保しようとします。スペインの援助を受けたマゼランが、南アメリカ大陸の南端を通過し、太平洋を横断します。マゼランはコロンブスとアメリゴベスプッチの後の世代だったので、これを成し遂げることができました。マゼランは、香辛料ですでに有名だったインドネシアのモルッカ諸島に行くつもりでしたが、途中のフィリピンに立ち寄ります。
これがフィリピンとスペインの縁の始まりです。
マゼランは、フィリピンでの戦闘で死亡してしまいます。マゼランの部下たちは、航海を続け、モルッカ諸島に行き、アフリカ大陸の南端を回って、スペインに戻りました。これが、初めての世界一周です。5隻あった船は1隻になり、270名いた船員は18名になるという、過酷な旅でした。
このマゼランの航海をきっかけに、スペインによるフィリピンの植民地化が始まりました。スペインによる植民地支配は、300年以上も続きました。
フィリピンは、スペインの王様のFelipe IIの島々として、Islas Filipinas(もとのeはiに変化)と呼ばれ、スペイン人によって統一が進められました。現在のThe Philippinesという国名は、ここから来ています。
スペインによる植民地支配の後には、アメリカによる植民地支配が待っていました。
参考文献
Lipson M. et al. 2014. Reconstructing Austronesian population history in Island Southeast Asia. Nature Communications 5(1): 4689.
Teixeira J.C. et al. 2021. Widespread Denisovan ancestry in Island Southeast Asia but no evidence of substantial super-archaic hominin admixture. Nature Ecology & Evolution 5(5): 616-624.