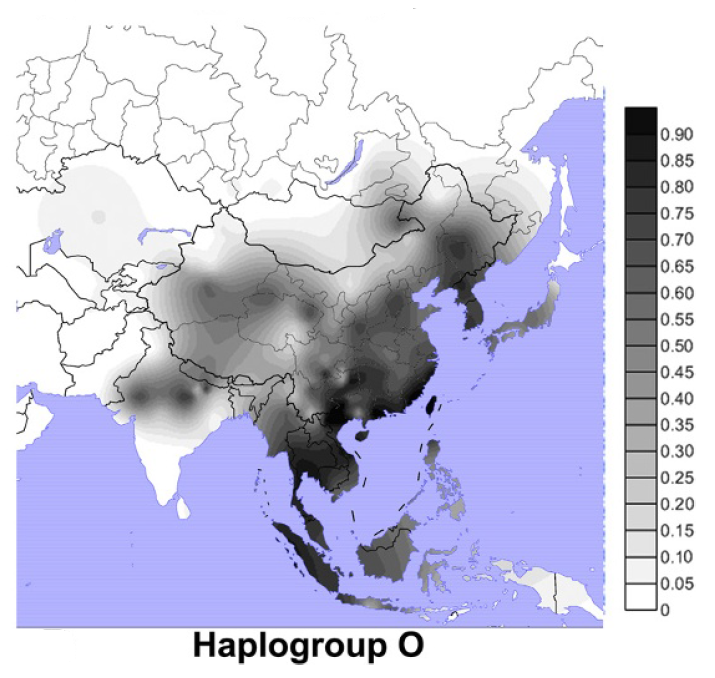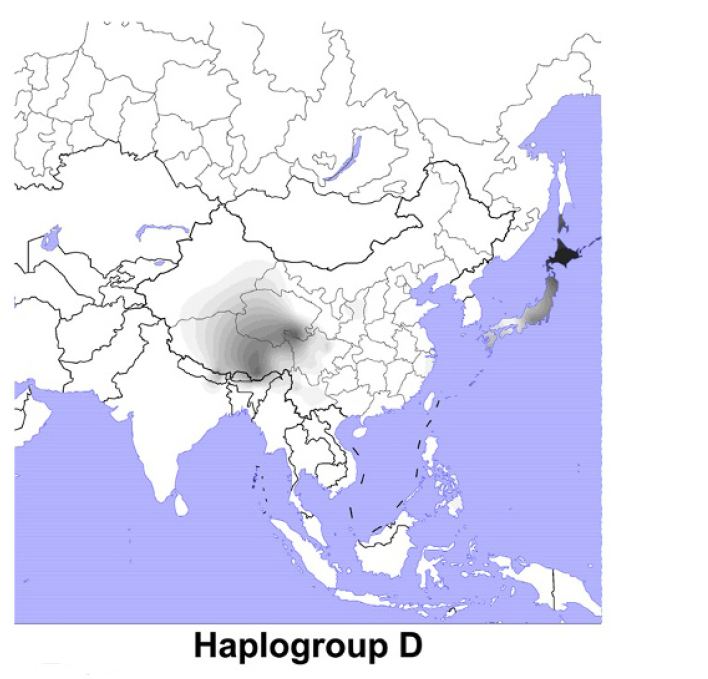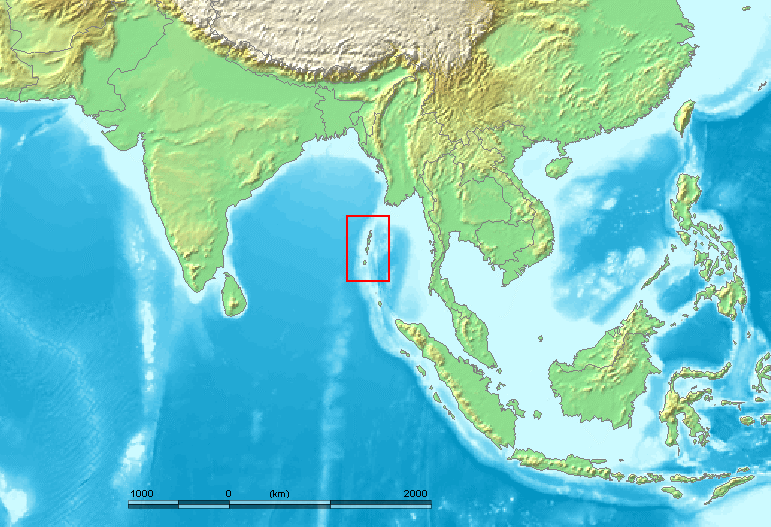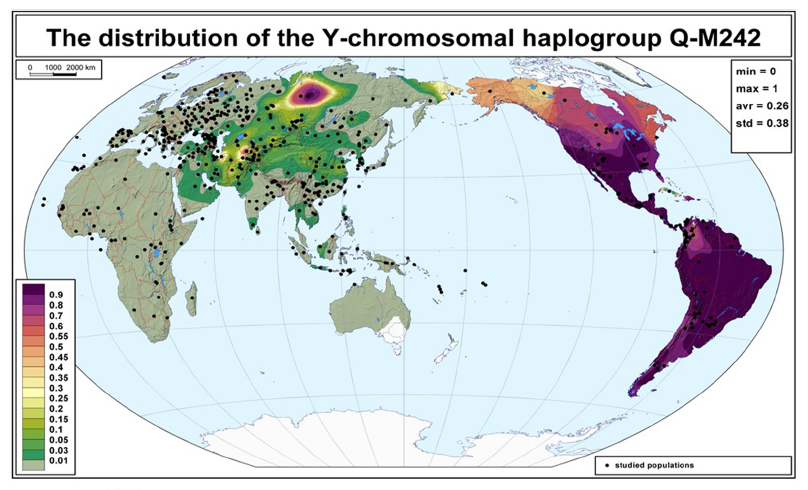

前回の記事では、アンダマン諸島が出てきました。アンダマン諸島なんて聞いたことがないという方も多いと思うので、少し説明しておきます(図はThangaraj 2003より引用)。
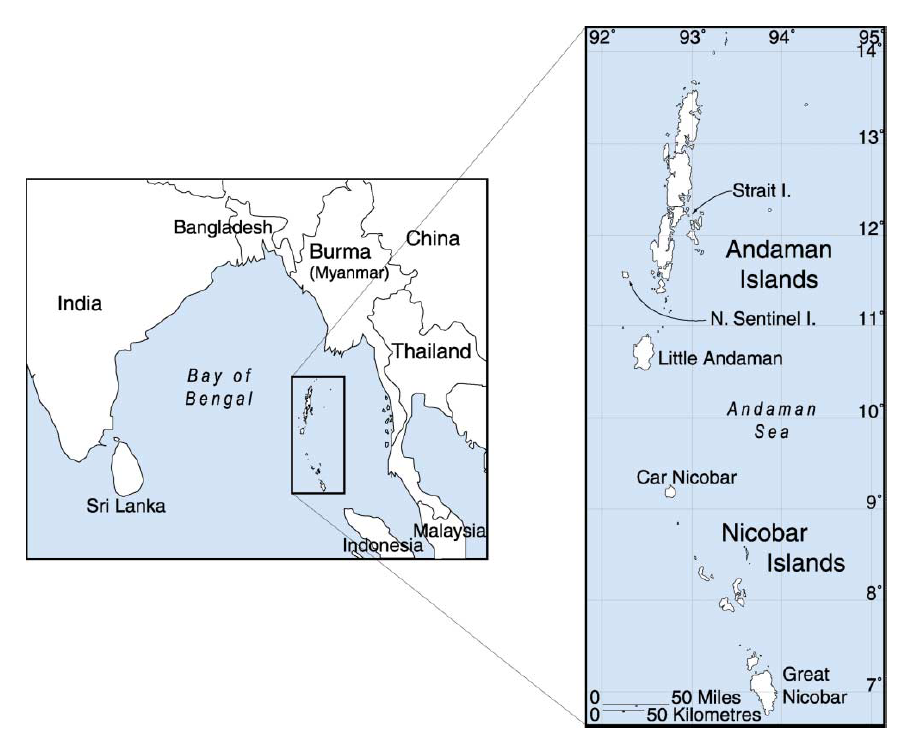
地図を見ればわかるように、アンダマン諸島はミャンマーの南方に位置し、現在ではインド領になっています。南アジアと東南アジアの境に近い微妙な位置です。
アンダマン人の外見は、南アジアの人々とも東南アジアの人々ともいくらか異なり、アフリカっぽいところもあります。アンダマン人の言語は、南アジアと東南アジアのどの言語にも似ていません。
ちなみに、アンダマン諸島の南にあるニコバル諸島には、典型的な東南アジアの人々が住み、ベトナム語やクメール語(カンボジアの主要言語)と類縁関係を持つ言語を話しています。
アンダマン人は、遠い昔にアフリカから南アジアあるいは東南アジアまでやって来て、その後孤立してしまった人たちではないかと考えられました。アフリカからオーストラリア・パプアニューギニアまで行った人々がいたが、その一部がアンダマン諸島に残ったのではないかと考えられました。アフリカ→中東→南アジア→東南アジア→オーストラリア・パプアニューギニアというルートはよく知られていたので、そのような考えが提示されたのは当然といえます。
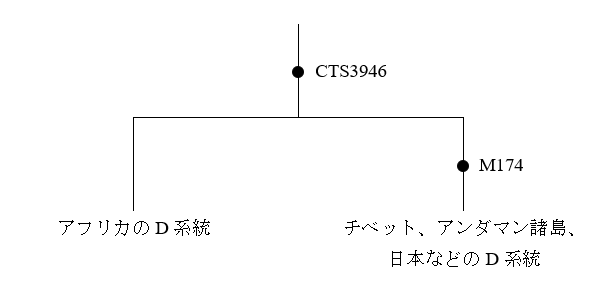
しかし、上の考えにはいろいろと問題があります。アンダマン諸島はごく小さな領域であり、アンダマン人のY染色体DNAおよびミトコンドリアDNAはほとんどバリエーションがありません。Y染色体DNAはD系統一色です。ミトコンドリアDNAはM31系統とM32系統の二つです。Y染色体DNAもミトコンドリアDNAもわけありです。
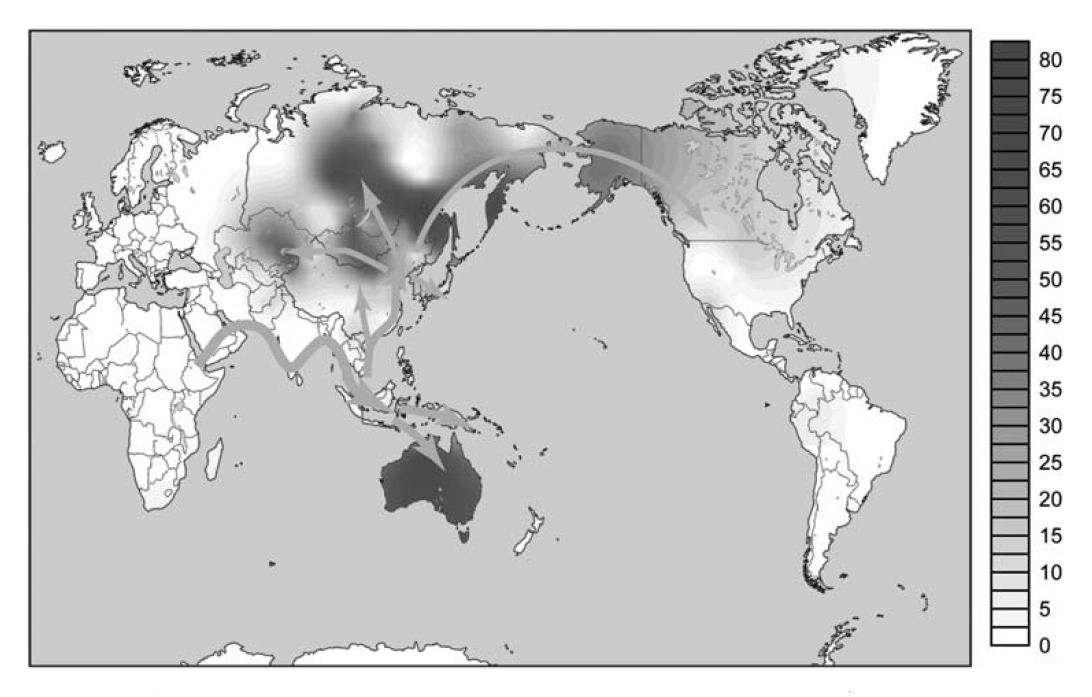
アンダマン人のY染色体DNAはD系統一色ですが、オーストラリア・パプアニューギニア方面にはD系統は全然見られません。オーストラリア・パプアニューギニア方面には、C系統とK系統は多く見られますが(K系統はF系統の一下位系統です)、D系統は全然見られないのです。アフリカからオーストラリア・パプアニューギニアに向かった人々の一部がアンダマン諸島に残ったというのは本当かと疑いたくなります。
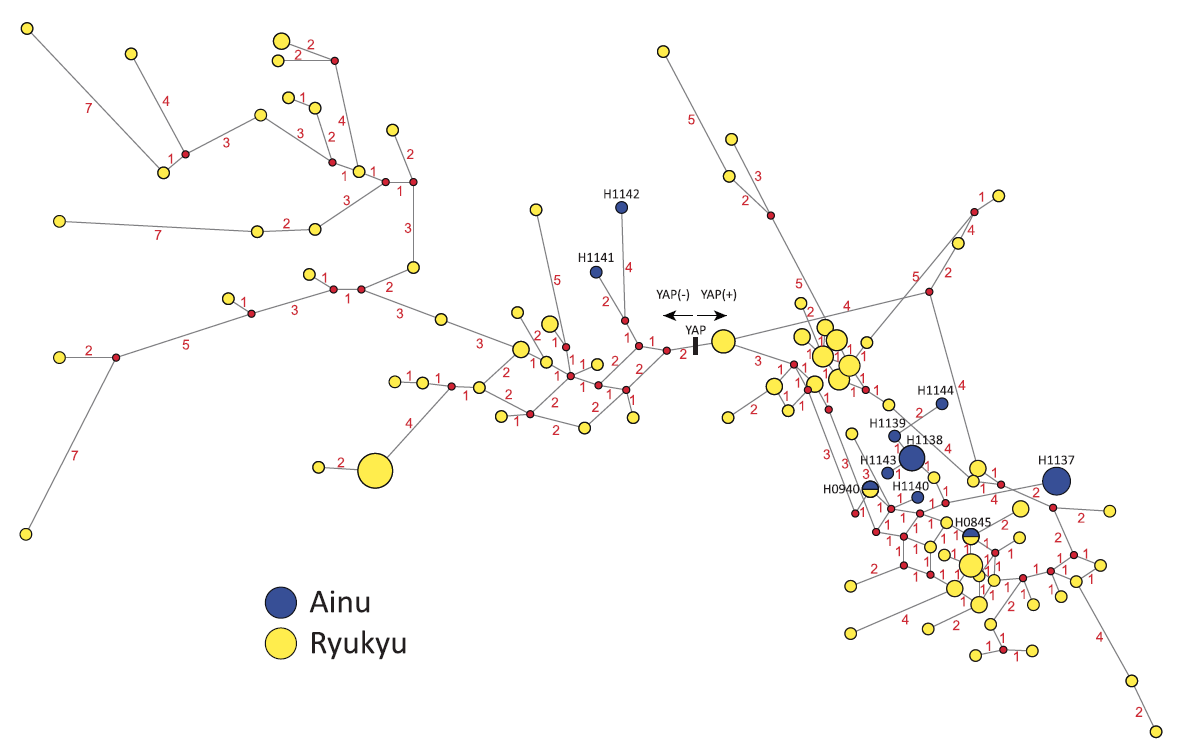
ミトコンドリアDNAのほうからも、そのような疑いを強める証拠が出てきました(図はWang 2011より引用)。
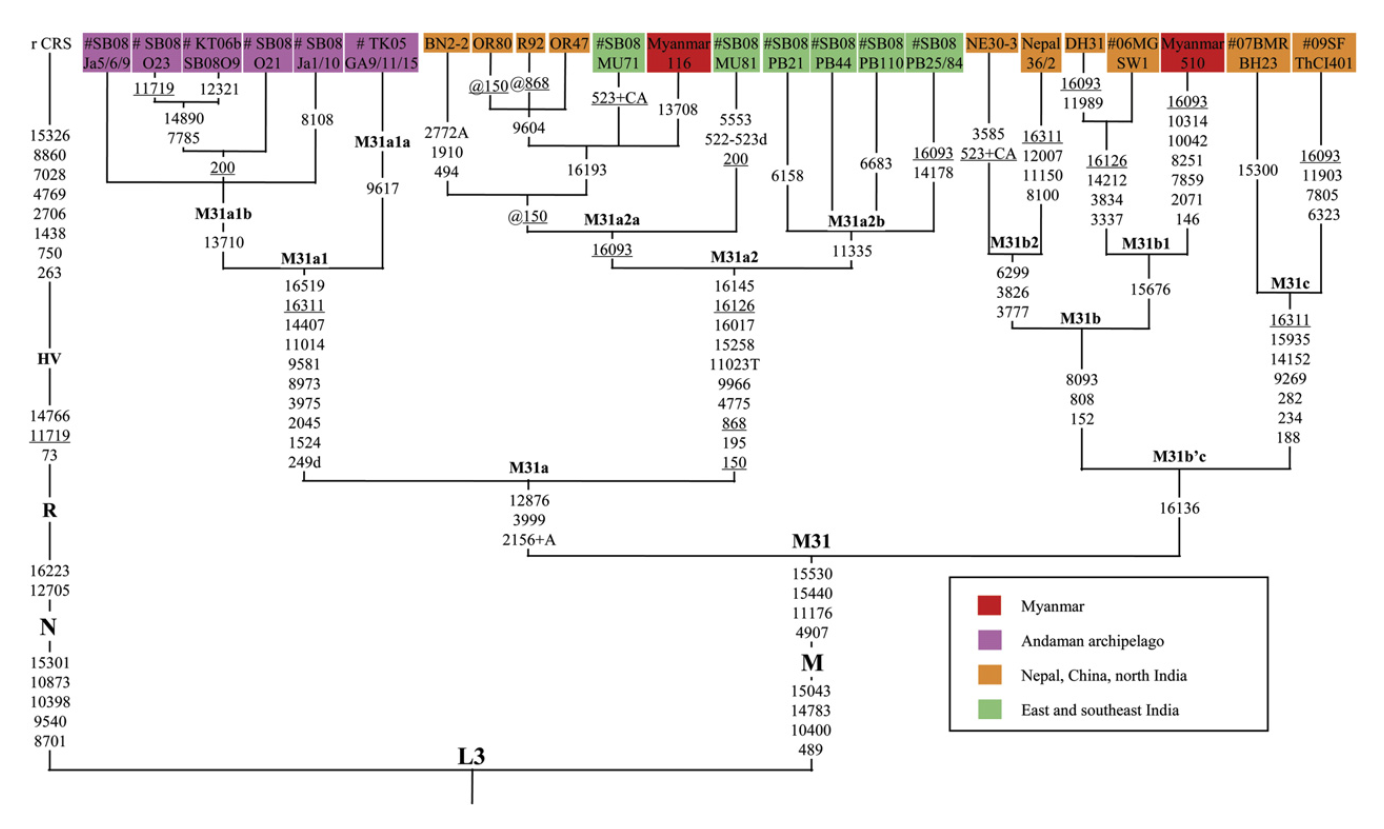
ミトコンドリアDNAのM系統は、アフリカの外で生じ、主に南アジアおよびそれより東の諸地域(東南アジア、オセアニア、東アジア、シベリア、アメリカ大陸)に広がりました。特に南アジアと東南アジアには、M系統の下位系統がたくさん存在し、その中にM31系統とM32系統があります。
M31系統は、アンダマン諸島だけでなく、インド、ネパール、ミャンマー、中国にも見られること、そしてM31系統の内部構造が上の図のようになっていることがわかってきました。アンダマン諸島のミトコンドリアDNAのタイプ(紫色)が、インド、ネパール、ミャンマー、中国のミトコンドリアDNAのタイプの中に埋め込まれているのがわかるでしょうか。
M32系統も、M31系統と大体同じ事情です。M系統の一下位系統としてM32’56系統があり、このM32’56系統がM32系統とM56系統に分かれています。M32系統はアンダマン諸島と東南アジアで見つかっており(Eng 2014)、M56系統は南アジアで見つかっています(Chandrasekar 2009)。
アンダマン人の先祖が南アジア・東南アジアの大陸部からアンダマン諸島に渡ったのは、人類が初めて南アジア・東南アジアに達した時代ではなく、いくらか後の時代である可能性が高くなってきました。総じて、M31系統もM32系統も大陸にほとんど残っておらず、アンダマン人の先祖が大陸を離れてから大陸が大きく様変わりしたことが窺えます。
●なぜY染色体DNAのD系統はオーストラリア・パプアニューギニア方面に見られないのか
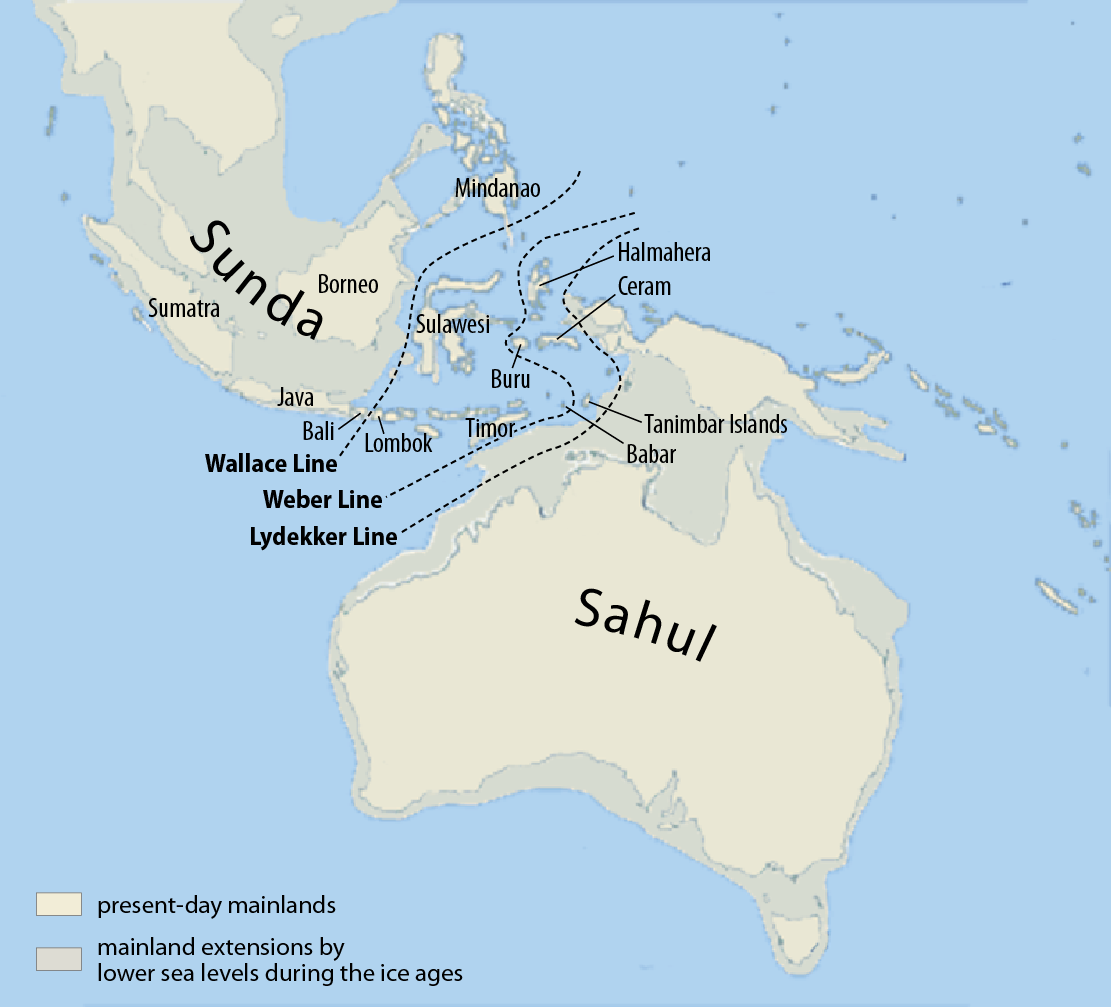
まず、東南アジアの地形が現在と大きく異なっていたことを思い出しましょう(図はScotese 2014より引用)。
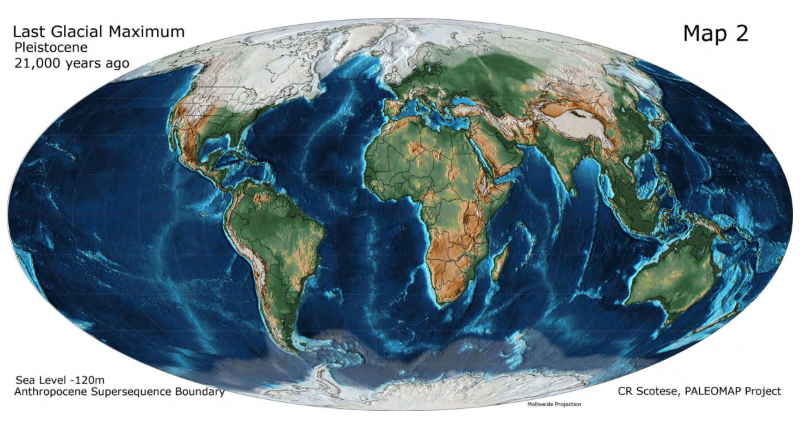
※この地図は、約2万年前のLast Glacial Maximum(最終氷期最盛期)の頃の世界地図です。人類がアフリカから東南アジアにやって来た5万年前頃は、もう少し海面が高いです。
東南アジアは、スンダランドという巨大な陸になっていました。その向かい側には、オーストラリアとパプアニューギニアがつながったサフルランドという巨大な陸がありました。スンダランドとフィリピンは、くっつきそうでくっついていなかったようです(Robles 2014)。
オーストラリア(アボリジニ)とパプアニューギニアには、Y染色体DNAのC系統とK系統が多く見られます。フィリピンのネグリトと呼ばれる原住民にも、C系統とK系統が多く見られます(Delfin 2011)。これらは、アフリカ→中東→南アジア→東南アジアと進んできた人の流れを示しています。スンダランドの海岸沿いを進み、スンダランドの先端からサフルランドとフィリピンに渡ったと考えられます。
ここで注目すべきなのは、Y染色体DNAのD系統がオーストラリア、パプアニューギニア、フィリピンに全くと言ってよいほど見られないということです(フィリピンでD系統の独自の下位系統がほんの数例見つかっているだけです(Y-DNA D Haplogroup Project – Y-DNA Classic Chart))。オーストラリア、パプアニューギニア、フィリピンでそうであるということは、スンダランドの海岸沿いを進んでスンダランドの先端に達した人間集団にD系統は全くあるいはほとんど存在しなかったと考えられそうです。
D系統がスンダランドの海岸沿いを進まなかったとすると、どのような展開が考えられるでしょうか。実は、大変重要な人骨の発見がありました(Demeter 2012)。この人骨は、ラオスのTam Pa Ling(タムパリン)で発見され、現生人類のものに間違いなく、少なくとも46000年前より古いと推定されました。50000~45000年前頃からオーストラリア・パプアニューギニア方面に人間の暮らしの形跡が現れ始めるので、それより前に人類が東南アジアに到達していたことは確実になっていましたが、それでも、ラオスのタムパリンの人骨の発見は驚きでした。なにが驚きだったかというと、海辺での生活からほど遠い内陸の奥深くから人骨が出てきたことです。
考えてみると、Y染色体DNAのD系統の豊かなバリエーションが見られるチベットのあたりはもろに内陸です。海辺とは正反対と言ってよい環境です。チベットのD系統(D-M15、D-P47、D*(まだよくわかっていないもの))、日本のD系統(D-M55)、アンダマン諸島のD系統(D-Y34637)は非常に古くに分かれており、最新の遺伝学の見積りでも50000年以上前に分かれたと推定されています(Mondal 2017)。D系統は50000年以上前に南アジアから東南アジアへの入口に来ていたのです。しかし、スンダランドの海岸に沿って南に進むことはしなかったのです。どうしたのでしょうか。先ほどの世界地図を見ればわかりますが、東南アジアの入口からスンダランドの海岸に沿って南に進むのも一つの道ですが、内陸を突っ切って東に進むのも一つの道です。ラオスのタムパリンで発見された人骨はまさに、内陸を突っ切って進む人々がいたことを示しています。D系統がこのような進路を取ったとすると、D系統がオーストラリア・パプアニューギニア方面に見られないことが納得できます。スンダランドの海岸を巡るのは大きな旅です。この大きな旅を経て東アジアにやって来た人々もいれば、この大きな旅を経ないで東アジアにやって来た人々もいたということです。アンダマン人の先祖は、南アジアから東南アジアへの入口の近くでしばらく過ごした後で、アンダマン諸島に渡ったのでしょう。
●もはや一本の流れでは説明できない東南アジアルート
4万年前の東アジアの記事などで、中東から中央アジア・シベリアを通って東アジアにやって来た人々と、中東から南アジア・東南アジアを通って東アジアにやって来た人々がいたことをお話ししてきました。要するに、中央アジアルートと東南アジアルート、あるいは北方ルートと南方ルートです。
人類が北方ルートと南方ルートの両方から東アジアにやって来たことがわかったのは、一つの前進です。しかし、北方ルートを単純な一本の流れ、南方ルートを単純な一本の流れと考えれば話は済むのかというと、そうもいきません。ここでは、南方ルートに焦点を当てましょう。
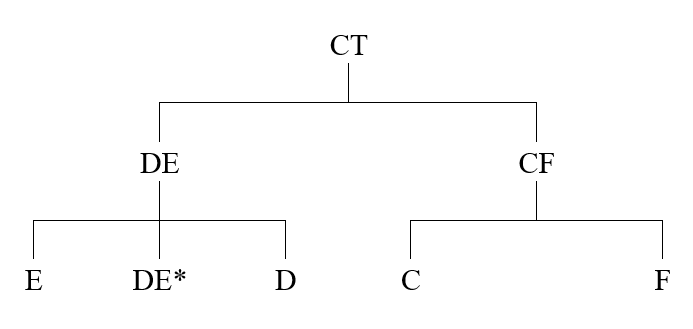
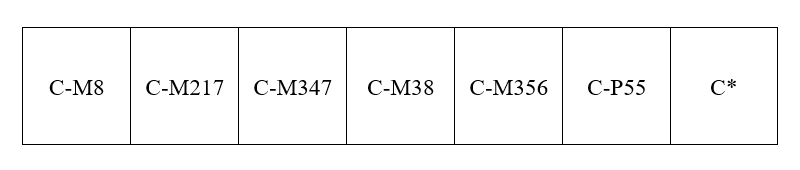
アフリカから始まる人類の歴史(Y染色体ハプログループCの研究)の記事で示しましたが、Y染色体DNAのC系統、D系統、F系統が東アジアの歴史に関係してきます。F系統は巨大な系統で、この系統からG系統、H系統、I系統、J系統、K系統が生まれました。F系統に属するが、G系統、H系統、I系統、J系統、K系統に向かう変異を起こしていない男性が少し残っており、それらの男性をF*と表します。
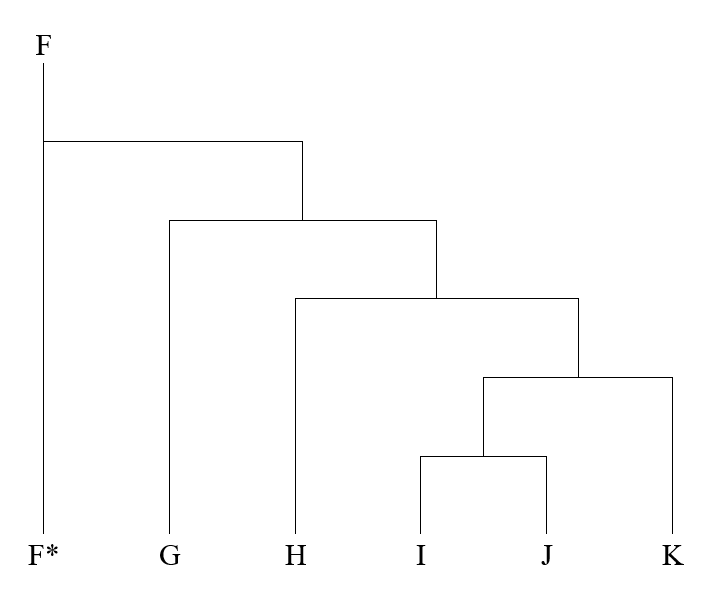
このうちのG系統、H系統、I系統、J系統は西ユーラシアにとどまり、K系統が東(オセアニア、東南アジア、東アジア、シベリア)に盛んに進出しました。F*グループは、現代では主に南アジアで見られますが、東南アジアと中国でもたまに見つかります。F*グループは東アジアにほんの少し進出した程度でしょう。
そのようなわけで、基本的に東アジアに進出してきたのは、C系統、D系統、K系統ということになります。オーストラリア・パプアニューギニア方面に、C系統とK系統は多く見られるが、D系統は全然見られないというのは、すでに述べた通りです。南アジアから東南アジアへの入口からスンダランドの海外沿いを進んだ人間集団にはD系統が全くあるいはほとんど存在せず、内陸に進んだ人間集団にはD系統が多かったわけです。このことを「一本の人の流れ」で説明するのは困難です。
アフリカ・中東→南アジア→東南アジアと進む一本の人の流れを想像してください。そこにはC系統もD系統もK系統も十分に存在します。東南アジアの入口に辿り着いたところで、一本の流れが二本の流れに分かれます。そのうちの一本の流れはほぼC系統とK系統だけで、もう一本の流れはD系統が支配的でした。確率はゼロではありませんが、考えづらいです。
海岸沿いを進むことに徹しながら東南アジアに至った人の流れと、海岸沿いを進むことにこだわらずに東南アジアに至った人の流れがあり、これらの流れの間の交流が(少なくともY染色体DNAの観点からは)限定的であったと考えたほうがはるかに自然です。C系統とK系統はCF系統から来ていますが、D系統はDE系統から来ており、発生場所にも隔たりがあったでしょう。C系統とK系統が互いに密に存在し、D系統が離れて存在していた可能性はあります。
南方ルートをもはや一本の人の流れで説明できないという問題が生じてきましたが、ほかにも考えなければならない問題があります。前々回と前回との記事でお話ししたように、Y染色体DNAのC系統とD系統は古くに東アジアに入っています。しかし、現在の東アジアではO系統が最大勢力です。そして、O系統と姉妹関係にあるN系統が北ユーラシアに大きく広がっています(Rootsi 2007の図を再掲)。
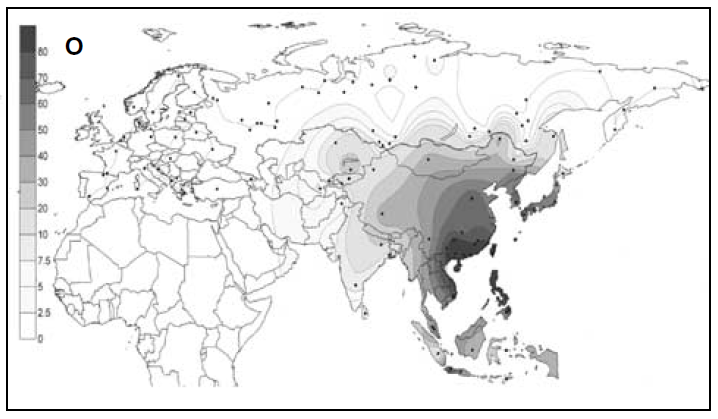
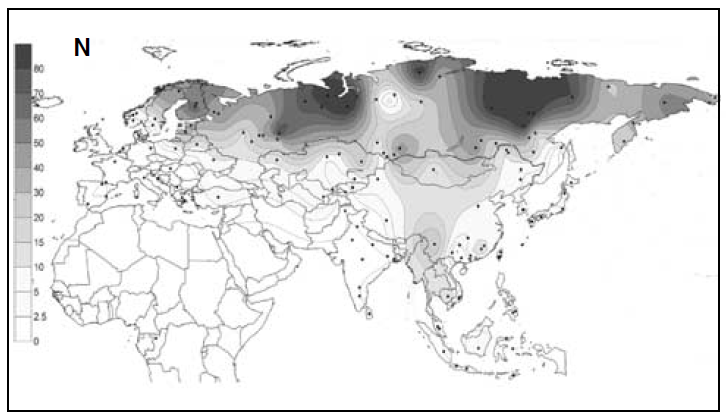
黄河文明と長江文明がO系統の勢力拡大と関係していること、遼河文明がN系統の勢力拡大と関係していることは間違いありません。東アジアの歴史を理解するには、O系統とN系統の理解が欠かせません。
ここで注意しなければならないのは、「O系統」と「N系統」という表記です。アフリカから始まる人類の歴史(Y染色体ハプログループCの研究)の記事で説明したように、Y染色体DNAのC系統、D系統、F系統がアフリカ以外の世界に広がっていきました。この中で、F系統は圧倒的に人数が多く、分布域が広いので、F系統の下位系統の記述には様々なアルファベット文字が用いられています。例えば、F系統の一下位系統がK系統で、K系統の一下位系統がNO系統で、NO系統の下位系統としてN系統とO系統があります。N系統とO系統の話というのは、K系統の内部の話、さらには、F系統の内部の話なのです。C系統とD系統に分類されるY染色体は歴史が深いが、N系統とO系統に分類されるY染色体は歴史が浅いと誤解してはいけません。N系統とO系統を見たら、K系統の問題だな、F系統の問題だなと認識しなければならないのです。アルファベット表記に惑わされないようにしましょう。
オーストラリア(アボリジニ)とパプアニューギニアにC系統に加えてK系統が見られるように、K系統も古くから東に進出しています。O系統とN系統は、その下位系統です。黄河文明、長江文明、遼河文明の起源と深く関係しますが、K系統のある下位系統がどのような過程を経て勢いづいたのか考えなければなりません。これもまた、東アジアの歴史における最重要問題の一つなのです。
参考文献
Chandrasekar A. et al. 2009. Updating phylogeny of mitochondrial DNA macrohaplogroup m in India: dispersal of modern human in South Asian corridor. PLoS One 4(10): e7447.
Delfin F. et al. 2011. The Y-chromosome landscape of the Philippines: Extensive heterogeneity and varying genetic affinities of Negrito and non-Negrito groups. European Journal of Human Genetics 19(2): 224-230.
Demeter F. et al. 2012. Anatomically modern human in Southeast Asia (Laos) by 46 ka. Proceedings of the National Academy of Sciences of the United States of America 109(36): 14375-14380.
Eng K. 2014. Complete mitochondrial DNA genome variation in Peninsular Malaysia. Doctoral dissertation, University of Leeds.
Mondal M. et al. 2017. Y-chromosomal sequences of diverse Indian populations and the ancestry of the Andamanese. Human Genetics 136(5): 499-510.
Robles E. et al. 2015. Late quaternary sea-level changes and the palaeohistory of Palawan Island, Philippines. Journal of Island and Coastal Archaeology 10(1): 76-96.
Rootsi S. et al. 2007. A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe. European Journal of Human Genetics 15(2): 204-211.
Scotese C. R. 2014. Atlas of Neogene Paleogeographic Maps (Mollweide Projection), Maps 1-7, Volume 1, The Cenozoic, PALEOMAP Atlas for ArcGIS, PALEOMAP Project, Evanston, IL.
Thangaraj K. et al. 2003. Genetic affinities of the Andaman Islanders, a vanishing human population. Current Biology 13(2): 86-93.
Wang H. et al. 2011. Mitochondrial DNA evidence supports northeast Indian origin of the aboriginal Andamanese in the Late Paleolithic. Journal of Genetics and Genomics 38(3): 117-122.