

人間のY染色体DNAが調べられるようになって年数が経過し、その分類も精緻になってきました。Y染色体DNAは、父から息子へ代々伝わるものです。日本ではY染色体DNAのO系統、D系統、C系統が多く見られますが、これらについて考察する前に、まずはすべての系統のおおもとであるアフリカのA系統に目を向けましょう(専門的には、ハプログループA、ハプログループC、ハプログループD、ハプログループOのように言いますが、本ブログでは一般の方にとって見慣れない専門用語を避けて、A系統、C系統、D系統、O系統のように言います)。
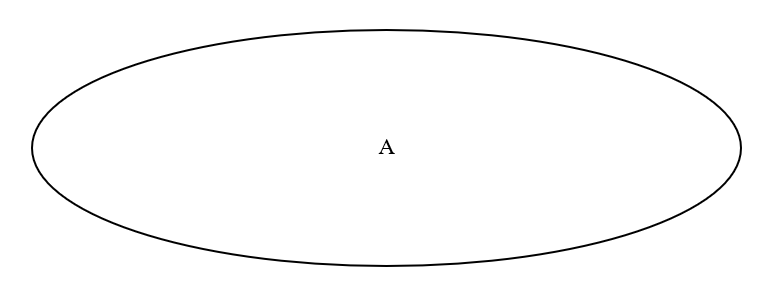
最初は、A系統の男性しかいませんでした(図はA系統の男性が集まっているところです)。
A系統のある男性に変異が起きて、BT系統が生まれます。
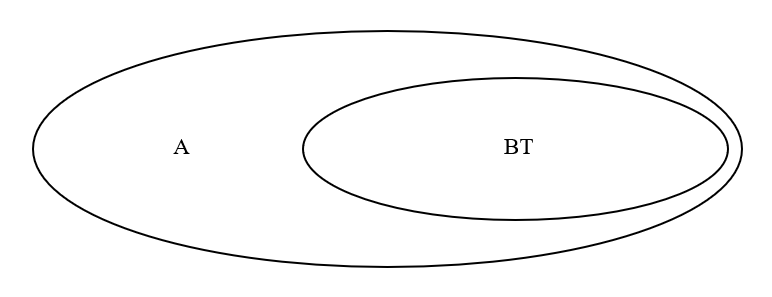
さらに、BT系統のある男性に変異が起きて、CT系統が生まれます。
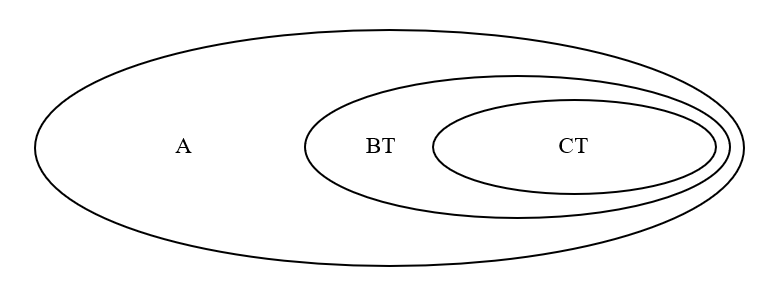
※正確に言うと、A系統の一下位系統からBT系統が生まれ、BT系統の一下位系統からCT系統が生まれましたが、ここでは単純に表現してあります。
ここまでは、アフリカで起きたことです。アフリカ以外のすべての男性のY染色体DNAは、このCT系統から来ています。CT系統から、複雑になっていきます。CT系統から、以下の系統が生まれました。
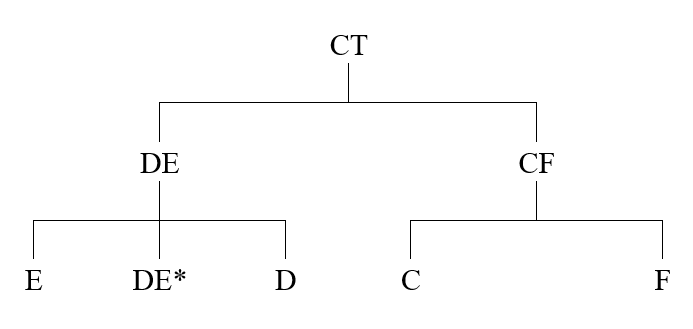
CT系統からDE系統とCF系統が生まれました。そして、DE系統からD系統とE系統が生まれ、CF系統からC系統とF系統が生まれました。
DE*という表記について説明しておきましょう。CT系統から変異を経てDE系統が生まれましたが、DE系統の男性の集まりを考えてください。

このうちの左端の男性にある変異が起き、D系統が生まれたとしましょう。そして、右端の男性に別の変異が起き、E系統が生まれたとしましょう。すると、D系統になるための変異も、E系統になるための変異も起きていない男性が残ります。実際に、そのような男性が少し残っているようなのです。そのような男性をDE*と書き表しています。
読者の方は、DE系統と同様のことがCF系統にもあったのではないかと考えるでしょう。もっともです。CT系統から変異を経てCF系統が生まれましたが、CF系統の男性の集まりを考えてください。

このうちの左端の男性にある変異が起き、C系統が生まれたとしましょう。そして、右端の男性に別の変異が起き、F系統が生まれたとしましょう。すると、C系統になるための変異も、F系統になるための変異も起きていない男性が残ります。実際に、そのような男性は存在したはずです。しかし、残っていないのです。そのため、上の系統図に「DE*」という表記はありますが、「CF*」という表記はないのです。
これまでに蓄積されているデータからすると、CT系統からDE系統への変異はアフリカで起きた可能性が高く、CT系統からCF系統への変異はアフリカの外で起きた可能性が高いです。アフリカにいたCT系統のある男性に変異が起きてDE系統が生まれ、アフリカの外にいたCT系統のある男性に変異が起きてCF系統が生まれたということです。
上の系統図のE系統、DE*グループ、D系統、C系統、F系統の分布について述べておきましょう。
E系統の分布は、圧倒的にアフリカに偏っています。ヨーロッパと中東に見られるE系統は、アフリカで生まれたE系統がかなり後になってアフリカの外に出たと考えられるものです。
F系統の分布は、圧倒的にアフリカの外に偏っています。アフリカに見られるF系統は、アフリカの外で生まれたF系統がかなり後になってアフリカに入ったと考えられるものです。アフリカの外の男性のY染色体DNAは、大部分がF系統です。ヨーロッパで支配的なR系統も、東アジア・東南アジアで支配的なO系統も、アメリカ大陸のインディアンで支配的なQ系統も、F系統の下位系統です。
DE*グループは、チベットで報告されていますが、報告例がごくわずかで、まだその存在が確実に認められていません(Shi 2008)。
残るは、C系統とD系統です。日本およびその周辺地域の遠い過去からの歴史を考えるうえで、まず重要になるのが、このC系統とD系統です。F系統と違って、C系統とD系統はアフリカの外で限定された特徴的な分布を示しています。その限定された特徴的な分布は、人類の歴史についてなにか物語っているようです。話の都合上、C系統から始めます。
●Y染色体DNAのC系統について
Y染色体DNAのC系統に関しては、H. Zhong氏らの優れた研究があります(Zhong 2010)。この研究は、Y染色体DNAのC系統が世界にどのように分布しているか調べ、C系統のかつての拡散経路を明らかにしたものです。Zhong氏らの結論を先に示します(図はZhong 2010より引用)。
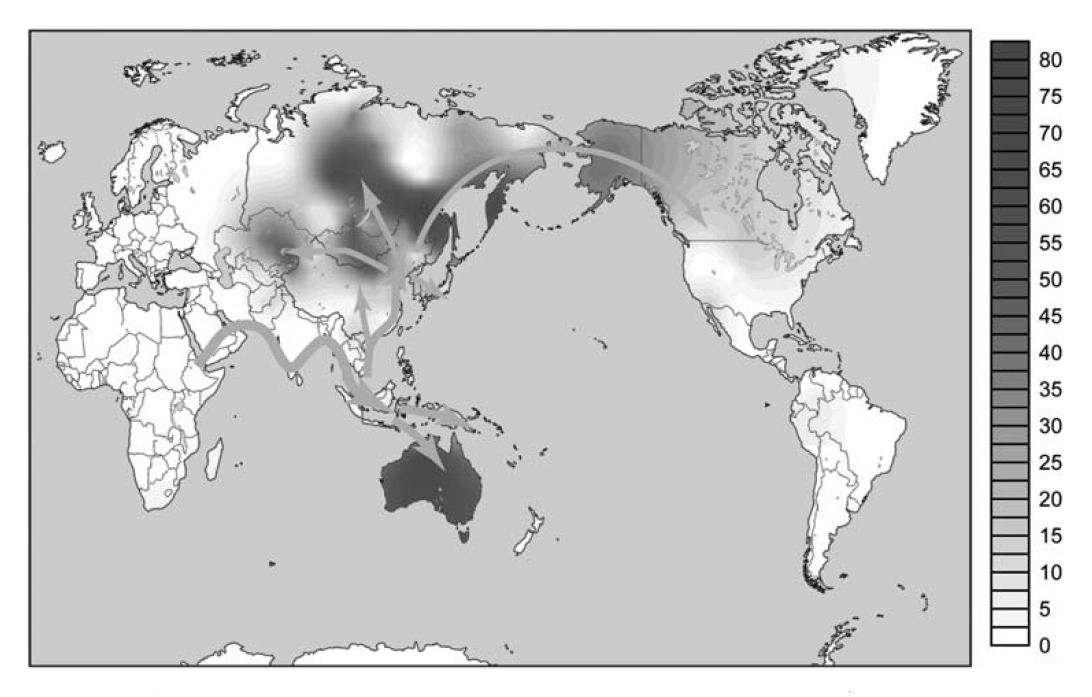
現在、Y染色体DNAのC系統は、以下の二つの地域で多く見られます。
・オーストラリアとその他のオセアニア地域
・東ユーラシアの北のほうを中心として、一方では中央アジアに、他方では北米北西部にかけて(東ユーラシアの北のほうにC系統がほとんど分布していないところがありますが、これはN系統が支配的なヤクート人のいるところです)
Zhong氏らは、C系統は中東から南アジアを通って東南アジアに進み、そこからオセアニア方面と東アジア方面に分かれたと考えています。Zhong氏らは、C系統の以下の下位系統の分布を調べています。
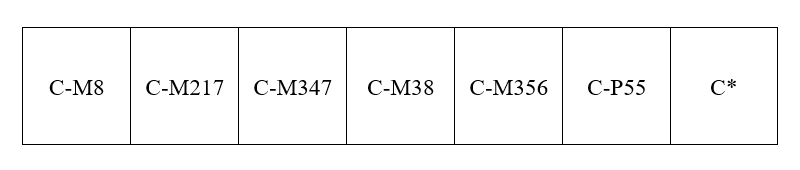
※Zhong氏らの論文では、旧表記が用いられています。C-M8=旧C1、C-M217=旧C3、C-M347=旧C4、C-M38=旧C2、C-M356=旧C5、C-P55=旧C6です。C*は、C-M8~C-P55のどれになる変異も起こしていないことを意味します。
Zhong氏らの研究は、2010年に発表されたものですが、DNAの研究・調査が急速に進んでおり、若干のアップデートが必要です。
上の下位系統の中でC-M347は、オーストラリア(アボリジニ)に非常に多く見られる系統です。C-M38は、オーストラリア以外のオセアニアに多く見られる系統です。上の下位系統の中でC-M347とC-M38は近い関係にありますが、両者の分岐はとても古いです。オーストラリアの住民とオーストラリア以外のオセアニアの住民が早くに分かれたことを示しています。
このC-M347とC-M38に系統上やや近いのが、C-M356です。C-M356は、インド周辺でわずかに見られる系統です。C-M347、C-M38、C-M356は、アフリカから中東に出て、中東→南アジア→東南アジアと進み、オーストラリアとそれ以外のオセアニアに分かれた人の流れを示しています。C-P55は、パプアニューギニアでわずかに報告されている系統です。
残るは、C-M8とC-M217です。実は、日本人に見られるC系統がこのC-M8とC-M217です。C-M8とC-M217を合わせて、日本人のY染色体DNAに占める割合は10%弱です(Hammer 2006)。
C-M217は、東ユーラシアの北のほう、中央アジア、北米北西部に大きく広がっている系統です。
C-M8は、日本以外でなかなか発見されず、謎めいていましたが、中国と朝鮮半島でわずかに見つかっています(www.yfull.com/tree/C/)。C-M8系統に関しては、驚くべき発見がありました。C-M8に系統上やや近いC-V20という系統がヨーロッパとその周辺にわずかに残っていることがわかってきました(Scozzari 2012)。中東でC系統のある下位系統が生じ、この下位系統が一方でヨーロッパ方面に、他方で東アジア方面に達したことを示唆しています。当然、その下位系統はヨーロッパと東アジアの間にも存在したはずです。激しい歴史展開の中で多くの系統が消滅し、遠い系統関係を持つ系統と系統が両極で見つかるよい例でしょう。
さて、C-M8とC-M217はどのようにして今の位置に到達したのでしょうか。先の地図のように、C系統が一方ではオセアニア方面、他方では東ユーラシアの北のほうで繁栄しているのを見ると、中東からの東南アジアルートと中央アジアルートを考えたくなります。しかし、これはいささか早計なようです。
まず、一夫一妻制ではない世界の記事の話を思い出してください。

中央アジアから東に進んできた人々と、東南アジアから北に進んできた人々が混ざり合った話です。●は中央アジアから東に進んできた男性、●は中央アジアから東に進んできた女性、■は東南アジアから北に進んできた男性、■は東南アジアから北に進んできた女性です。
中央アジアからやって来た男性が力関係(権力・武力)あるいは物質的豊かさの点で東南アジアからやって来た男性より優位にあったため、上の図のような子作りが行われ、中央アジアからやって来たY染色体DNAが残ることになりました。
しかし、上の図をよく見てください。中央アジアからやって来た人数と東南アジアからやって来た人数を比べれば、東南アジアからやって来た人数のほうが断然多いのです。■の男性、すなわち東南アジアからやって来た男性は子作りに参加していませんが、これらの男性はどのようなタイプのY染色体DNAを持っていたのでしょうか。東南アジアからやって来たわけですから、そのY染色体DNAはQ系統ではありえません。Q系統は、●の男性、すなわち中央アジアからやって来た男性が持っていたY染色体DNAです。
東南アジアから北に進んできた■の男性のY染色体DNAは、中央アジアから東に進んできた●の男性のY染色体DNAに遮られて、それ以上北に進むことができません。つまり、東南アジアから北にある程度進んだものの、そこで止まってしまったY染色体DNAの系統があったのです。このY染色体DNAは何系統だったのでしょうか(もちろん、画一的だったとは限りません)。
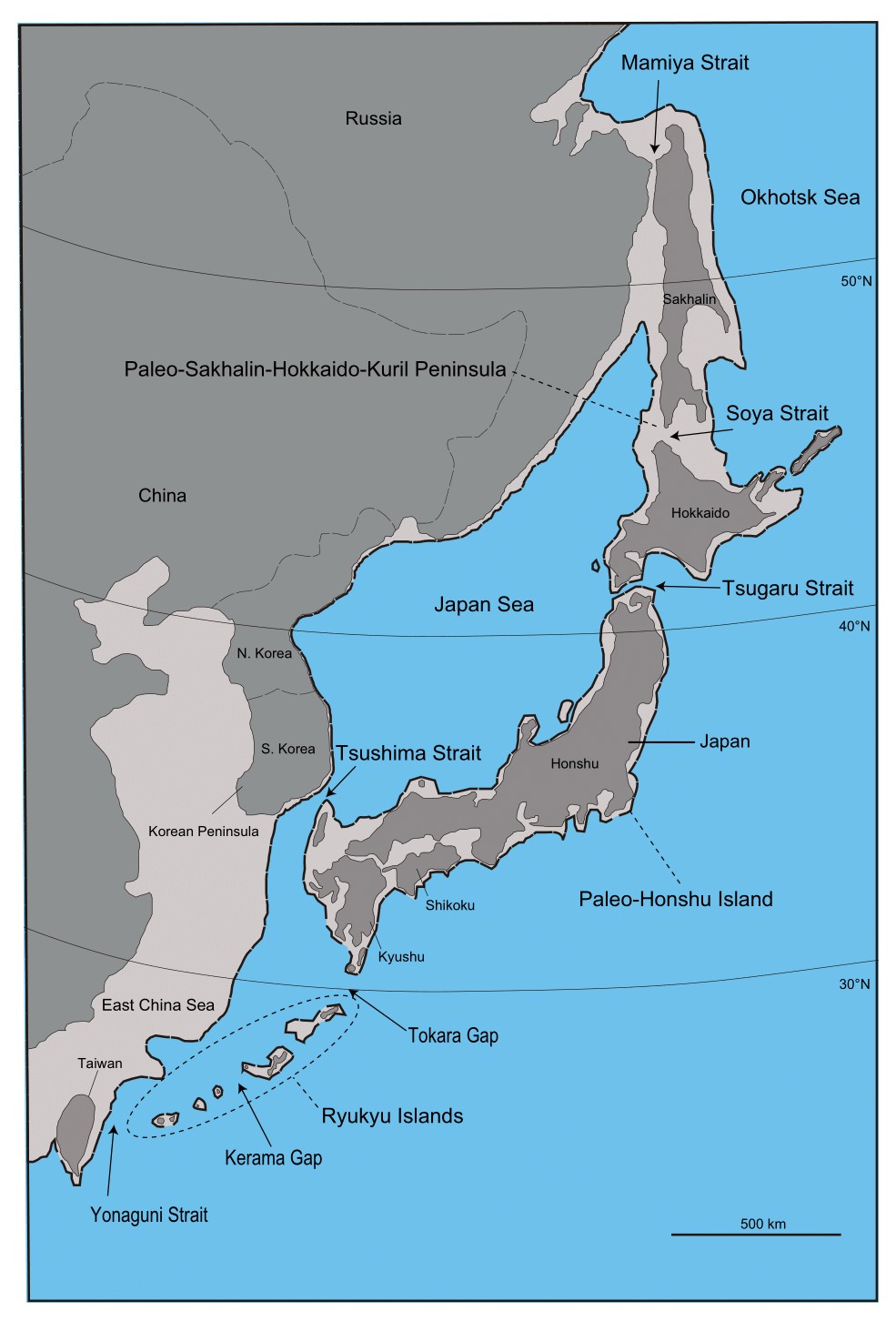
次に、「日本列島は大陸と陸続きだった」という言い方には注意が必要の記事の話を思い出してください(図はWikipediaの図を再掲)。
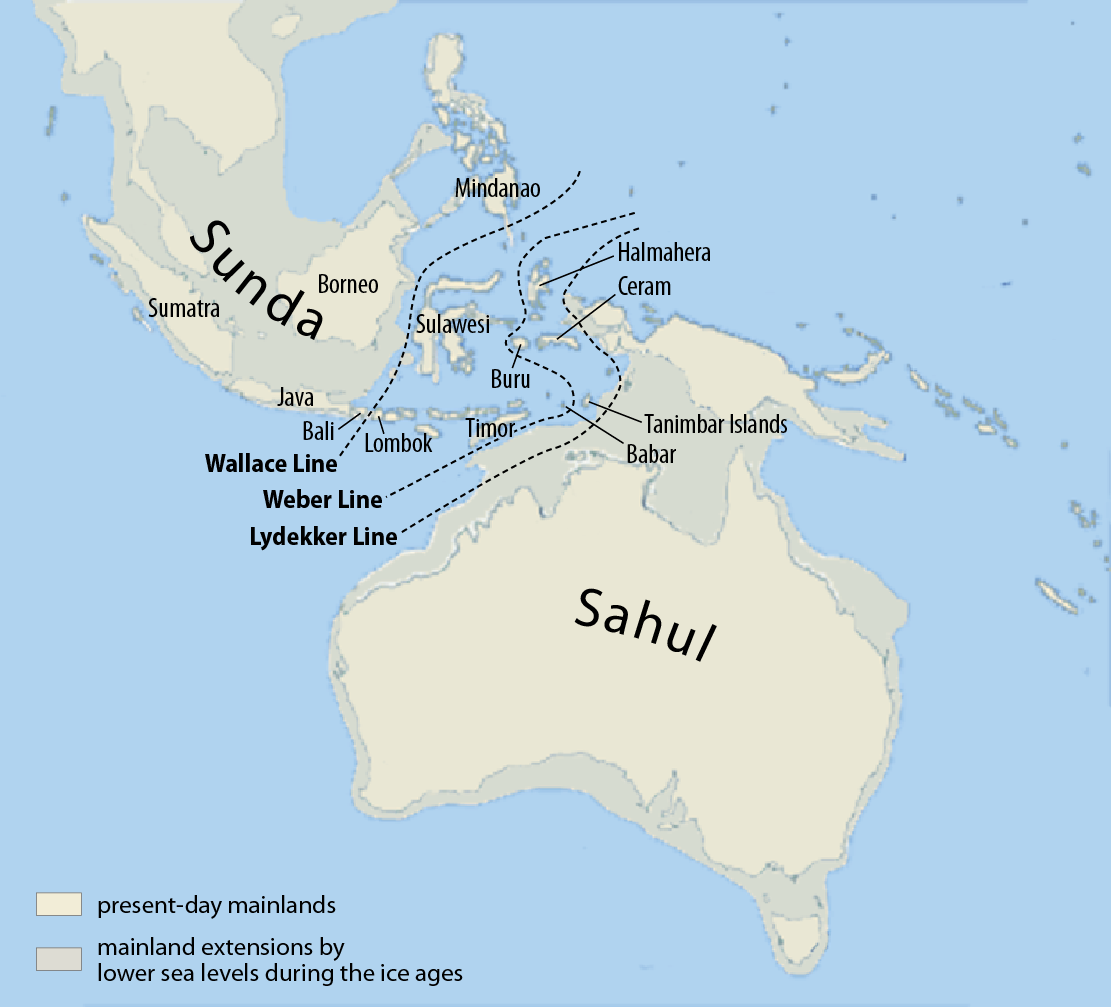
アフリカから中東に出て、中東から南アジアを通って東南アジアに達する人の流れがありました。この人の流れは、海岸沿いを進んでいく人の流れです。当時の東南アジアはスンダランドという巨大な陸になっていたというのが重要なポイントです。
上の地図の左上(南アジア方面)から人がやって来て、海岸沿いを進んでいきます。スンダランドからサフルランドに渡るのも一つの道ですが、さらに海岸沿いを進んで右上(東アジア方面)に抜けていくのも一つの道です。当時の東南アジアの地形を考えると、オセアニア方面に向かうか、東アジア方面に向かうかというのは、ほんのわずかな選択の差だったのです。オセアニア方面で高い航海能力を示した人々と、東アジア方面で高い航海能力を示した人々の話をしましたが、これらの人々の出所は同じである可能性が高いです。こうなると、オセアニア方面と東アジア方面に共通しているY染色体DNAの系統が大きな関心になります。そしてオセアニア方面と東アジア方面に共通しているY染色体DNAの系統というのが、まさにC系統なのです。
●アメリカ大陸のインディアンに見られるY染色体DNAのQ系統とC系統について再び考える
アメリカ大陸のインディアンに特徴的なY染色体DNAのQ系統の分布図を再び示します(Balanovsky 2017の図を再掲)。
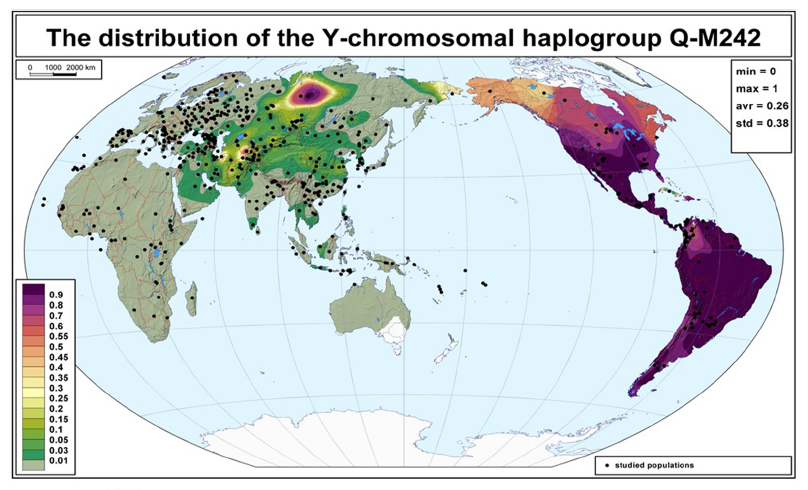
南米のインディアンのY染色体DNAがほぼQ系統一色であることはお話ししました。しかし、南米のインディアンのY染色体DNAにも、わずかにC系統が存在します(Mezzavilla 2014)。このC系統は、C-M217に属します。しかし、ユーラシアおよび北米で知られているC-M217の各下位系統に当てはまりません。そのため、アメリカ大陸に最初に入っていった人々のY染色体DNAはほぼQ系統一色であったが、わずかにC系統(C-M217、ただし現在ユーラシアと北米で知られているタイプとは少し異なるタイプ)が存在していたのではないかと推測されています。
アメリカ大陸に最初に入っていった人々は、約2万年前のLast Glacial Maximum(最終氷期最盛期)の前にシベリアにいた人々です。この人々のY染色体DNAはほぼQ系統一色で、わずかにC系統が存在していたのではないかと思われます。LGMより前にC系統がシベリアに入ろうとしても、Q系統が大きな存在感を誇っていたわけです。南米のインディアンのY染色体DNAはそのことを示しています。その後、LGMが始まって、シベリアはほぼ住めない状態になり、ベーリング地方の人間集団と東アジアの人間集団に二分されます。LGMが終わり、16000年前頃から、北米を覆っていた氷が解け、ベーリング地方の人間集団はアメリカ大陸に入っていきます(このあたりの事情については、閉ざされていたアメリカ大陸への道およびベーリング陸橋、危ない橋を渡った人々を参照)。と同時に、ほぼ住めない状態になっていたシベリアも再び住める状態になります。LGM前のシベリアではQ系統が支配的であったが、いったん人がいなくなり、LGM後のシベリアにはC系統が突進していったと見られます。そう考えると、北米北西部でQ系統が大きく減り、C系統が大きく増えていることが理解できます。アメリカ大陸に最初に入っていった人々にはC系統はほとんど存在しなかったが、アメリカ大陸に後から入っていった人々にはC系統が多かったということです。
LGMが終わり、再び住めるようになったシベリアには、C系統だけでなく、N系統も進出していきました(変わりゆくシベリアを参照)。しかし、C系統は北米北西部に及んでいますが、N系統は北米北西部に及んでいません。C系統が東アジアから北上し始めた時期は、N系統が東アジアから北上し始めた時期より早かったのかもしれません。C系統はLGMが終わってすぐに東アジアから北上し始め、N系統はしばらく経って遼河文明が起こる頃に東アジアから北上し始めたのではないかということです。
C-M217は東ユーラシアの北のほうに大きく広がっていますが、C-M217の内部のバリエーションは、中央アジアの側では乏しく、中国東部、朝鮮、日本の側では豊かです(理解するためにはShort Tandem Repeatの知識も必要であり、これについては次回の記事で説明します)(Zhong 2010)。C-M217は、中央アジアのほうから中国東部のほうに広がったのではなく、中国東部のほうから中央アジアのほうに広がったと考えられます。中国東部のほうに存在したC-M217の一部が中央アジアのほうに広がったということです。
以上すべてを総合すると、以下のように言えそうです。
Y染色体DNAのC系統は中東から南アジアを通って東南アジアに達し、そこからオセアニア方面と東アジア方面に分かれた。東アジアにはC-M8やC-M217などのいくつかの下位系統が到達したが、中央アジアからやって来た寒冷地や内陸での暮らしに長けた男性が力関係(権力・武力)あるいは物質的豊かさの点で優位にあり、C-M8やC-M217はそれ以上北に進むことがほとんどできなかった。C-M8は、東アジア(主に日本)に少数派として残った。C-M217は、LGM前はシベリアにほとんど進出することができなかったが、LGM後はシベリアに盛んに進出し、中央アジアと北米北西部にも至った。
東アジアの歴史の根本を考えるうえで、C系統と並んで重要なのがD系統です。C系統の分布は独特ですが、D系統の分布も独特です。D系統の話に移りましょう。
参考文献
Balanovsky O. et al. 2017. Phylogeography of human Y-chromosome haplogroup Q3-L275 from an academic/citizen science collaboration. BMC Evolutionary Biology 17: 18.
Hammer M. F. et al. 2006. Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes. Journal of Human Genetics 51(1): 47-58.
Mezzavilla M. et al. 2014. Insights into the origin of rare haplogroup C3* Y chromosomes in South America from high-density autosomal SNP genotyping. Forensic Science International: Genetics 15: 115-120.
Scozzari R. et al. 2012. Molecular dissection of the basal clades in the human Y chromosome phylogenetic tree. PLoS ONE 7(11): e49170.
Shi H. et al. 2008. Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations. BMC Biology 6(1): 45.
Zhong H. et al. 2010. Global distribution of Y-chromosome haplogroup C reveals the prehistoric migration routes of African exodus and early settlement in East Asia. Journal of Human Genetics 55(7): 428-435.