

ミトコンドリアDNAとY染色体DNAだけでなく、DNA全体が調べられるようになり、興味深い事実が明らかになってきました。その中でまず注目すべきなのは、現生人類が古人類(現生人類に近い種)と交わっていたという事実です。現生人類と古人類の問題は、例えば日本語の起源や日本語の系統には直接関係ありませんが、現生人類の思考と言語(今の私たちの思考と言語のようなもの)がいつ頃形成されたのかを考えるうえで、無視することができません。
現生人類が6万年前頃にアフリカを出て、ユーラシアに広がっていったことは本ブログでお話ししていますが、現生人類に近い種は、それよりはるかに前からユーラシアに広がっていました。皆さんも「北京原人」や「ジャワ原人」のような名前を聞いたことがあるでしょう。「北京原人」は80万年前頃から今の中国北部に、「ジャワ原人」は160万年前頃から今のインドネシアにいたことがわかっています(Ciochon 2009)。ちなみに、「北京原人」と「ジャワ原人」と呼ばれていましたが、両者に特に大きな違いはなく、今ではホモ・エレクトスという一つの種にまとめられています。ホモ・サピエンスはラテン語で「賢い人」、ホモ・エレクトスはラテン語で「直立した人」という意味です。
実は、現生人類に近い種の発見は今も続いており、インドネシアのフローレス島で新しい種が発見されたり(Brown 2004)、フィリピンのルソン島で新しい種が発見されたりしています(Détroit 2019)。どうやら、現生人類が進出する前のユーラシアには、現生人類に近い様々な種が存在したようです。しかし、現生人類がユーラシアに進出するにつれて、現生人類以外の種はユーラシアから姿を消していきました。なにがあったのだろうと考えたくなるところです。
古人類の中で脳容量と骨格が現生人類に近く、現生人類との関係が注目されてきたのが、ネアンデルタール人です。この名前は、ドイツのネアンデル谷(ドイツ語で谷はTal)から来ています。現生人類とネアンデルタール人の間に交雑(異種間で子を作ること)があったかどうかということについては、論争が繰り広げられていましたが、現生人類のDNA全体とネアンデルタール人のDNA全体が調べられるようになった現在では、はっきり答えが出ています。答えはイエスです。しかし、ミトコンドリアDNAとY染色体DNAを調べてもわからず、DNA全体を調べて明らかになりました。
現生人類の多くの人々のミトコンドリアDNAとY染色体DNAが調べられてきましたが、ネアンデルタール人のミトコンドリアDNAまたはY染色体DNAの系統が見つかったという報告はありません。やはり、ミトコンドリアDNAは、女→女→女→女→女という形でしか伝わらないので、なかなか子孫に残らないのです。どこまでも女でなければならないわけですから、大変厳しいです。同様に、Y染色体DNAも、男→男→男→男→男という形でしか伝わらないので、なかなか子孫に残らないのです。どこまでも男でなければならないわけですから、大変厳しいです。
そこで、前回の記事でお話しした第1染色体~第22染色体とX染色体の出番です。現生人類の男性とネアンデルタール人の女性の間で子が作られたとしましょう(現生人類がネアンデルタール人に対して優位であったことを考えると、現生人類の男性と現生人類の女性の間で子が作られ、さらに、現生人類の男性とネアンデルタール人の女性の間で子が作られる傾向があったと推測されます)。その子は、以下の染色体を持つことになります。
女の子なら・・・

男の子なら・・・
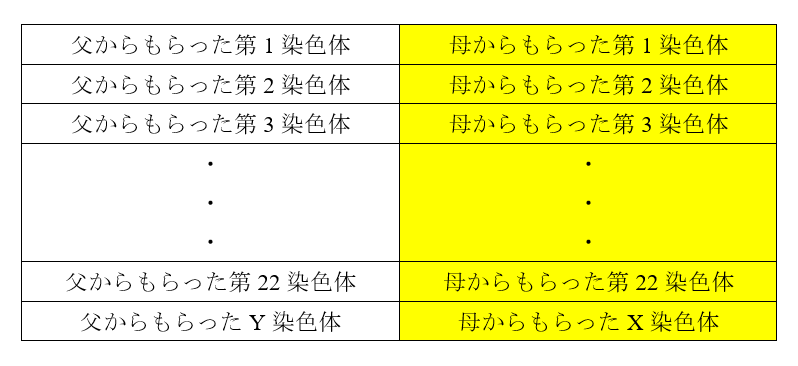
ネアンデルタール人の母からもらった第1染色体~第22染色体およびX染色体にあるDNA配列は、前回の記事で説明したように、時に切断されながら、一部が子孫に残されていきます。
ネアンデルタール人の母からもらった第1染色体はとてつもなく長いので、そこにある特定の短いDNA配列は、切断を免れて、そのまま子孫に残る可能性があります(「そのまま」と書きましたが、長い時間が経過すれば変異も起きるので、「ほぼそのまま」が正確です)。
第2染色体にある特定の短いDNA配列、・・・、第22染色体にある特定の短いDNA配列、X染色体にある特定の短いDNA配列も、切断を免れて、そのまま子孫に残る可能性があります。
そのようなわけで、ネアンデルタール人の母からもらったいずれかの染色体にあった特定の短いDNA配列が、切断を免れて、そのまま遠い遠い子孫に残る可能性は十分にあります。
実際に、近年のバイオテクノロジーの目覚ましい進歩によって、ネアンデルタール人のDNAを調べ、現生人類のDNAと比較することができるようになりました。そしてまもなく、現生人類の常染色体(第1染色体~第22染色体)とX染色体に、ネアンデルタール人から来たと考えられる部分が見つかり始めました(Green 2010、Yotova 2011、Vernot 2014)。
ネアンデルタール人が現生人類と交わったことを証明するのだから、ネアンデルタール人のDNA配列と現生人類のDNA配列に共通部分を見つけるのかなと、皆さんは予想するのではないでしょうか。確かに、その通りです。しかし、注意しなければならないことがあります。以下のような構図を考えてください。
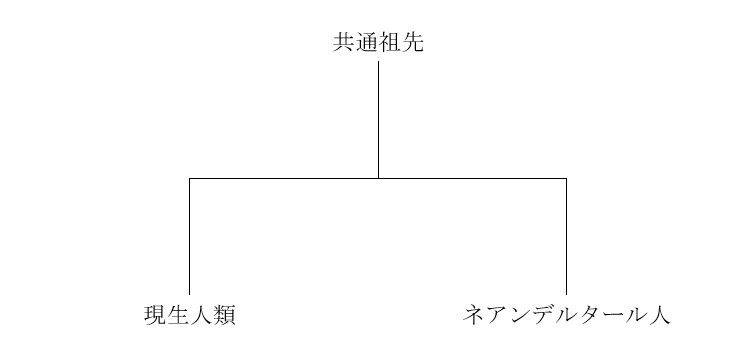
単にネアンデルタール人のDNA配列と現生人類のDNA配列に共通部分を見つけただけでは、「その共通部分は共通祖先から来たのだろう」と言われた時に、反論するのが困難です。ネアンデルタール人が現生人類と交わったことを証明するためには、どうすればよいでしょうか。
当初から主張されていましたが、決め手になったのは、「アフリカの現生人類」のDNA配列と「アフリカの外の現生人類」のDNA配列に明確な差があるということでした(Green 2010、Yotova 2011)。具体的に言うと、「アフリカの外の現生人類」のDNA配列が、ところどころで、「アフリカの現生人類」のDNA配列ではなく、ネアンデルタール人のDNA配列と共通性を示していたのです。そこから、以下のような構図が浮かび上がってきます。
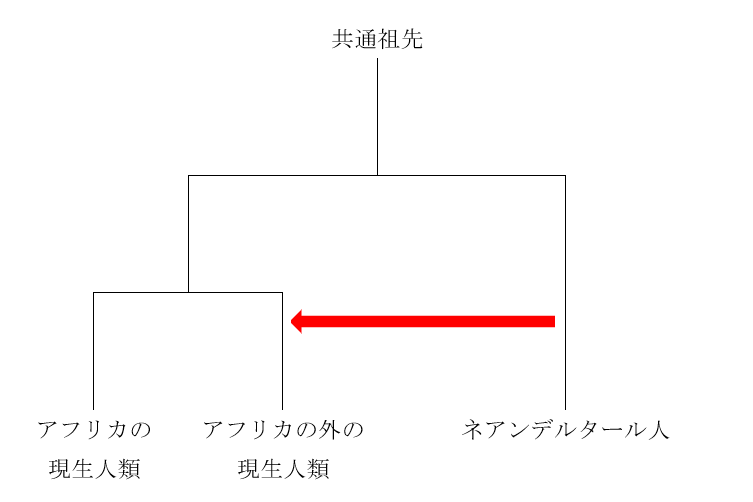
図中の赤い矢印は、ネアンデルタール人が「アフリカの外の現生人類」と交わっているところです。ネアンデルタール人由来の部分は「アフリカの外の現生人類」に広く見られるため、現生人類はアフリカを出てすぐに、つまり方々に散らばっていく前に、ネアンデルタール人と交わったと考えられています(Green 2010、Yotova 2011)。
現代の「アフリカの外の現生人類」のDNAに占めるネアンデルタール人由来の部分の割合は大きくなく、数パーセント以下です(ユーラシアからアフリカに戻った人々もいたため、アフリカでも完全にゼロではありません)(Prüfer 2014、Sankararaman 2014)。ちなみに、ヨーロッパでネアンデルタール人の遺骨・遺跡がいくつも見つかっているので、ネアンデルタール人というとヨーロッパのイメージがありますが、ヨーロッパの人々より東アジアの人々でネアンデルタール人由来の部分の割合が若干大きいこともわかってきました(Vernot 2015)。「アフリカから出る→すぐにネアンデルタール人と交わる→方々に散らばる」だけでは十分に説明しきれていないことを示唆しており、東アジア方面でネアンデルタール人の要素が追加された可能性や、ヨーロッパ方面でネアンデルタール人の要素が薄められた可能性が検討されています(Vernot 2015)。
このように、現生人類がネアンデルタール人と交わっていたことを示す証拠が出され始めましたが、ちょうどその頃、別のところで大きな事件が起きていました。
2008年に、ロシアのアルタイ地方のデニソワ洞窟で謎の人骨が発見されました。まずミトコンドリアDNAが調べられ、骨の主は現生人類とネアンデルタール人と共通祖先を持つ未知の種であることが明らかになりました(Krause 2010)。
参考文献
Brown P. et al. 2004. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431(7012): 1055-1061.
Ciochon R. L. et al. 2009. Asian Homo erectus converges in time. Nature 458(7235): 153-154.
Détroit F. et al. 2019. A new species of Homo from the Late Pleistocene of the Philippines. Nature 568(7751): 181-186.
Green R. E. et al. 2010. A draft sequence of the Neanderthal genome. Science 328(5979): 710-722.
Krause J. et al. 2010. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature 464(7290): 894-897.
Prüfer K. et al. 2014. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505(7481): 43-49.
Sankararaman S. et al. 2014. The genomic landscape of Neanderthal ancestry in present-day humans. Nature 507(7492): 354-357.
Vernot B. et al. 2014. Resurrecting surviving Neandertal lineages from modern human genomes. Science 343(6174): 1017-1021.
Vernot B. et al. 2015. Complex history of admixture between modern humans and Neandertals. American Journal of Human Genetics 96(3): 448-453.
Yotova V. et al. 2011. An X-linked haplotype of Neandertal origin is present among all non-African populations. Molecular Biology and Evolution 28(7): 1957-1962.