
ベーリング地方の近くに位置するYana RHS遺跡で発見された二人の男性のミトコンドリアDNAはU系統で、Y染色体DNAはP系統であることが判明しました(Sikora 2019)。
※女性だけでなく男性も自分の母親からもらったミトコンドリアDNAを持っています。ただし、男性の場合は、そのミトコンドリアDNAを子どもに伝えることができません。
ミトコンドリアDNAのU系統も、Y染色体DNAのP系統も、日本ではなじみがありませんが、人類の歴史を考える際には非常に重要な系統です。まずは、ミトコンドリアDNAのU系統に注目しましょう。
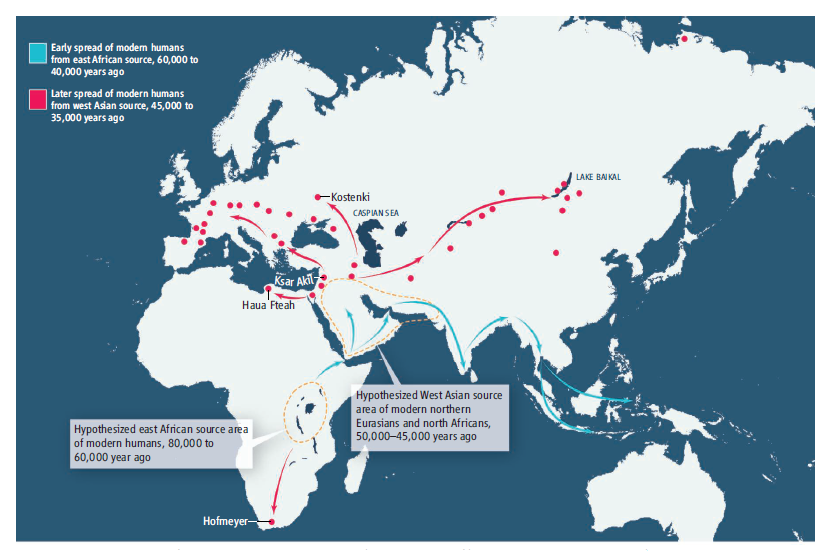
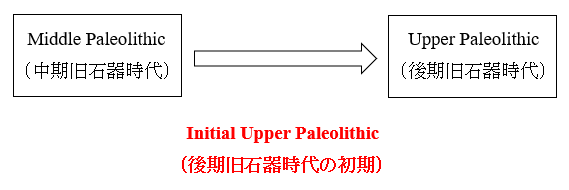
前回の記事では、4.5~5万年前頃から中東のレバント地方(今のシリア、レバノン、ヨルダン、イスラエルのあたり)でInitial Upper Paleolithic(後期旧石器時代の初期)と呼ばれる動きが始まり、この先進的な動きがヨーロッパとアフリカ北東部に拡散したことをお話ししました。
現在のヨーロッパではミトコンドリアDNAのH系統の割合が最も高くなっていますが(Achilli 2004)、人類が進出してまもない頃のヨーロッパではミトコンドリアDNAのU系統が支配的でした(Soares 2010、Posth 2016)。
中東のレバント地方からヨーロッパとアフリカ北東部にInitial Upper Paleolithic(後期旧石器時代の初期)を拡散させた人々の主なミトコンドリアDNAがU系統であったと考えられています(Maca-Meyer 2001、Olivieri 2006)。
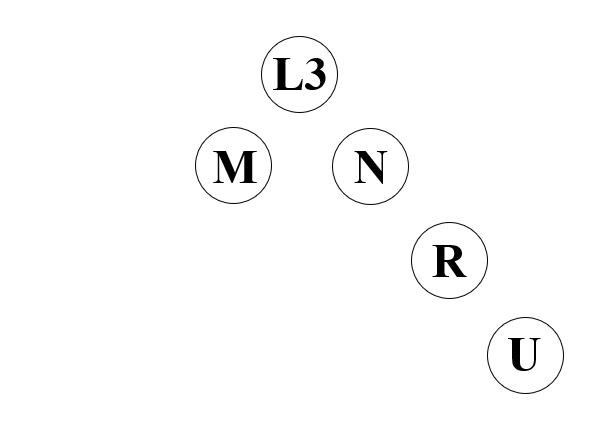
※アフリカに見られるミトコンドリアDNAのL3系統から、アフリカの外に見られるM系統とN系統が生まれました。そして、N系統の一下位系統としてR系統が生まれ、R系統の一下位系統としてU系統が生まれました。
4.5~5万年前頃に中東のレバント地方で支配的だったミトコンドリアDNAのU系統がすぐにヨーロッパとアフリカ北東部に入り込んだのはわかりますが、そのU系統が遅くとも3万年前頃にユーラシア大陸のほぼ最北東部といってよいYana RHS遺跡に到達していたというのは驚きです。
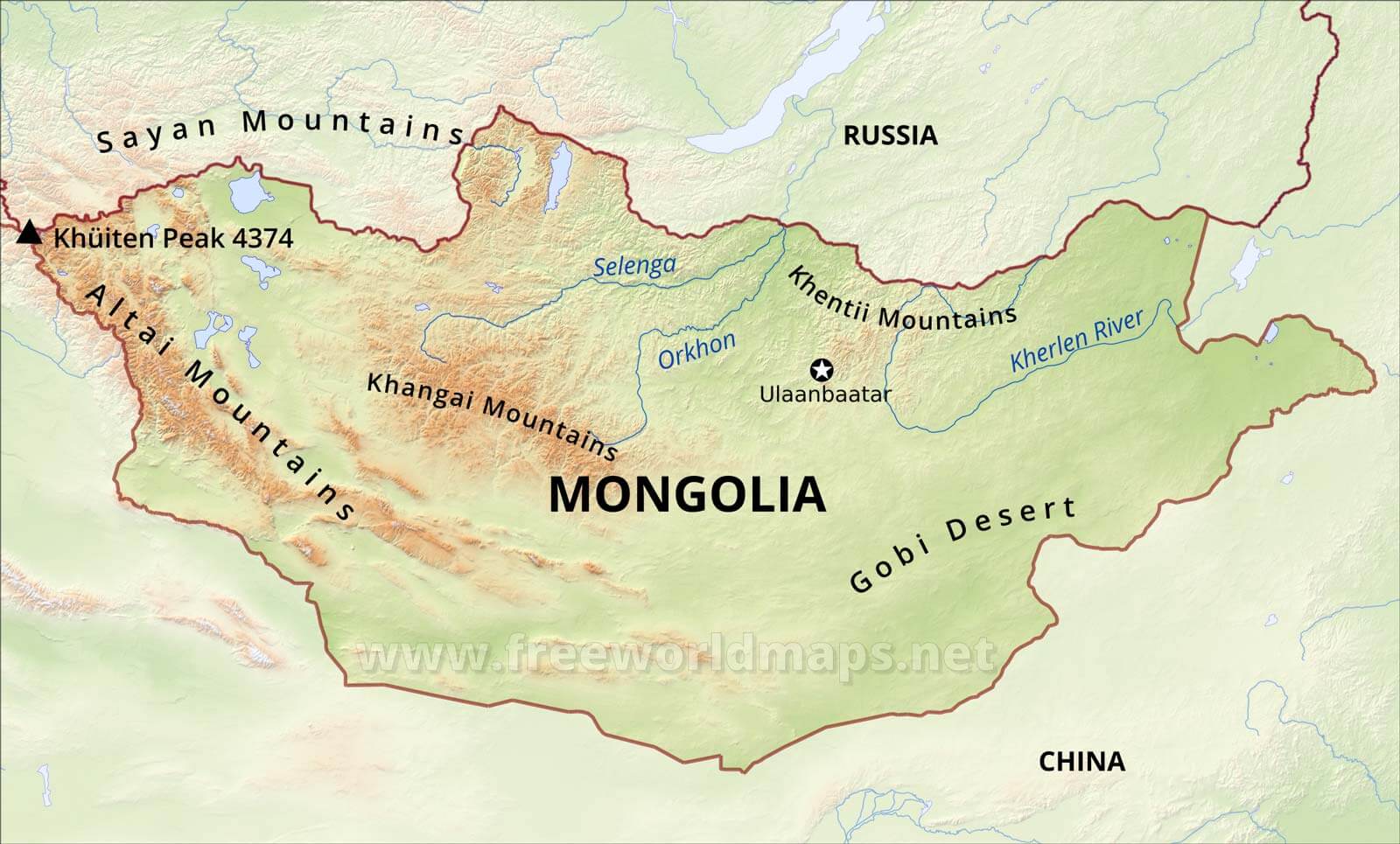
しかし、この発見は、バイカル湖の近くに位置するロシアのMal’ta(マリタ)で発見された2.4万年前の男の子のミトコンドリアDNAがU系統であったこと(Raghavan 2014)、そして中央アジアの北のほうに位置するロシアのUst’-Ishim(ウスチイシム)で発見された4.5万年前の男性のミトコンドリアDNAがR系統(つまりU系統の一段階前のタイプ)であったこと(Fu 2014)とよく合います。
ちなみに、日本の近辺で発見された現生人類として最も古いのは、以前に言及した北京郊外の田園洞遺跡で発見された4万年前の男性ですが、この男性のミトコンドリアDNAはB系統でした(Fu 2013)。上で説明したように、U系統はR系統の一下位系統ですが、B系統もR系統の一下位系統です。このことは注意を引きます。しかし、もとのR系統は人類がアフリカを出て早い段階で発生しており、パプアニューギニア・オーストラリアに見られるP系統もR系統の一下位系統です。そのため、田園洞の男性のミトコンドリアDNAのB系統が中央アジアのほうから来たのか、東南アジアのほうから来たのかという問題は慎重に検討する必要があります。
これまでに発見されている古代北ユーラシアの人間の骨または歯はわずかですが、ミトコンドリアDNAのN系統、R系統およびその下位系統が中東から北ユーラシアに広がっていく様子が見えそうです。
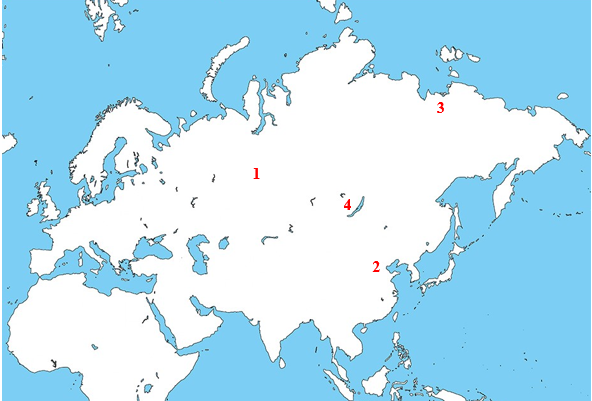
1 Ust’-Ishim 2 田園洞 3 Yana RHS 4 Mal’ta
ミトコンドリアDNAだけでなく、Y染色体DNAにも一貫性が感じられます。Ust’-Ishimの男性のY染色体DNAはK系統(Fu 2014)、田園洞の男性のY染色体DNAは発表されていませんが、Yana RHSの二人の男性のY染色体DNAはP系統(Sikora 2019)、Mal’taの男の子のY染色体DNAはR系統でした(Raghavan 2014)。K系統の一下位系統としてP系統があり、P系統の下位系統としてアメリカ大陸のインディアンで支配的なQ系統とMal’taの男の子のR系統があります。もととなったK系統はI系統とJ系統と近縁で、I系統はヨーロッパを中心に分布し、J系統は中東を中心に分布しています(Rootsi 2004、Semino 2004)。やはり、中東から北ユーラシア全体への拡散があったことを窺わせます。遺跡の多いアルタイ山脈周辺とバイカル湖周辺は、その拡散において重要な役割を果たしたのでしょう。
※上記の五人の古代人男性のほかに、バイカル湖のいくらか右下に位置するモンゴルのSalkhit(サルヒト)でも、性別不明ながらN系統のミトコンドリアDNAを持つ3.4万年前の人物が発見されています(Devièse 2019)。
こうなると気になるのは、中東→中央アジア→バイカル湖周辺と拡散してきたInitial Upper Paleolithic(後期旧石器時代の初期)の担い手たちは、中国、朝鮮、日本方面にどのような影響を与えたのかということです。前回の記事でお話ししたように、中東で始まった先進的なInitial Upper Paleolithic(後期旧石器時代の初期)は40000~43000年前頃にはバイカル湖周辺に到達しているので、ちょうどその頃、そしてその少し後の東アジアでなにが起きていたか調べなければなりません。4万年前頃の東アジアはどのようになっていたのでしょうか。
※現在のヨーロッパを大きく支配しているのはY染色体DNAのR系統(R1a系統とR1b系統)で、インド・ヨーロッパ語族はこのR系統と深い関係があるのではないかと注目されてきました(Manco 2015)。上に示した古代北ユーラシアの人々のY染色体DNAを見ると、R系統とインド・ヨーロッパ語族の根源がヨーロッパからかなり離れたところにありそうなことがわかります。Mal’taの男の子のY染色体DNAは、R1a、R1bあるいはこれらの共通祖先であるR1に分類されるための変異を起こしておらず、R系統の中で原初的なタイプです(Raghavan 2014)。
参考文献
Achilli A. et al. 2004. The molecular dissection of mtDNA haplogroup H confirms that the Franco-Cantabrian glacial refuge was a major source for the European gene pool. American Journal of Human Genetics 75(5): 910-918.
Devièse T. et al. 2019. Compound-specific radiocarbon dating and mitochondrial DNA analysis of the Pleistocene hominin from Salkhit Mongolia. Nature Communications 10(1): 274.
Fu Q. et al. 2013. DNA analysis of an early modern human from Tianyuan Cave, China. Proceedings of the National Academy of Sciences of the United States of America 110(6): 2223-2227.
Fu Q. et al. 2014. Genome sequence of a 45,000-year-pld modern human from western Siberia. Nature 514(7523): 445-449.
Maca-Meyer N. et al. 2001. Major genomic mitochondrial lineages delineate early human expansions. BMC Genetics 2: 13.
Manco J. 2015. Ancestral Journeys: The Peopling of Europe from the First Venturers to the Vikings. Thames & Hudson.
Olivieri A. et al. 2006. The mtDNA legacy of the Levantine early Upper Palaeolithic in Africa. Science 314(5806): 1767-1770.
Posth C. et al. 2016. Pleistocene mitochondrial genomes suggest a single major dispersal of non-Africans and a Late Glacial population turnover in Europe. Current Biology 26(6): 827-833.
Raghavan M. et al. 2014. Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans. Nature 505(7481): 87-91.
Rootsi S. et al. 2004. Phylogeography of Y-chromosome haplogroup I reveals distinct domains of prehistoric gene flow in Europe. American Journal of Human Genetics 75(1): 128-137.
Semino O. et al. 2004. Origin, diffusion, and differentiation of Y-chromosome haplogroups E and J: Inferences on the neolithization of Europe and later migratory events in the Mediterranean area. American Journal of Human Genetics 74(5): 1023-1034.
Sikora M. et al. 2019. The population history of northeastern Siberia since the Pleistocene. Nature 570(7760): 182-188.
Soares P. et al. 2010. The Archaeogenetics of Europe. Current Biology 20(4): R174-R183.