

●Y染色体DNAのD系統について
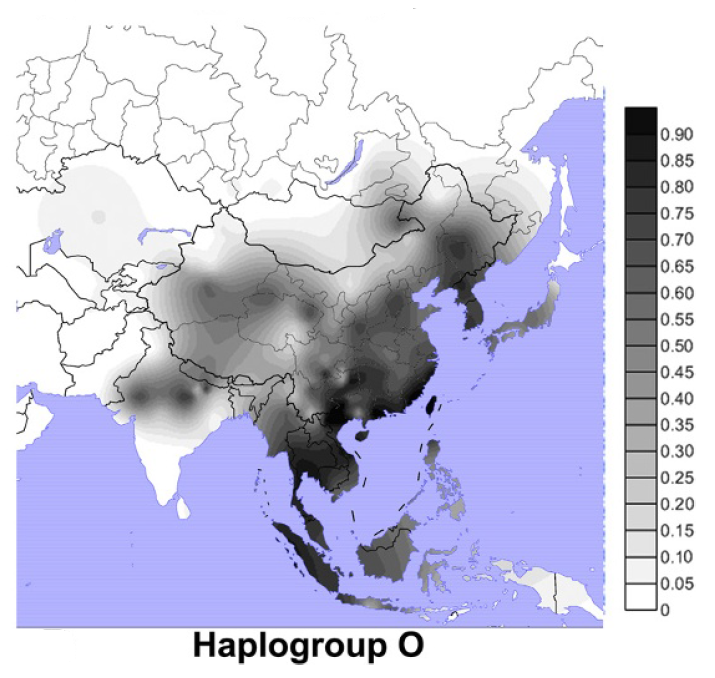
Y染色体DNAのD系統(Y染色体ハプログループD)は、日本人のDNAの話をする時に盛んに注目されてきた系統です。日本全体では、O系統が一番多く、D系統が二番目に多くなっています(Hammer 2006ではO系統51.8%、D系統34.7%、Nonaka 2007ではO系統54.4%、D系統39.2%)。
D系統が注目されてきたのは、O系統が日本の近隣地域によく見られるのに対して、D系統が日本の近隣地域にほとんど見られないからです(図はWang 2013より引用)。
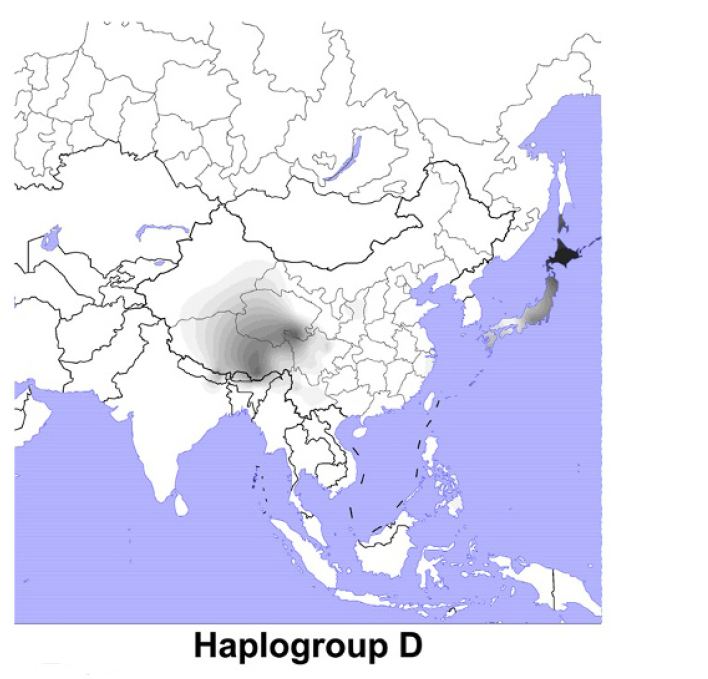
D系統の分布は独特です。チベット側と日本側に分かれています。昔の人々は歩いて移動していたわけですから、かつてはチベットと日本の間の領域にもD系統がたくさん存在したはずです。D系統が支配的だった空間が、O系統が支配的な空間に変わったことが窺えます。農耕の誕生・発展(黄河文明と長江文明)が大きく影響したことは間違いないでしょう。
ちなみに、O系統とD系統の割合は日本内部でも地域差があります(Hammer 2006、Nonaka 2007)。明らかにO系統が優勢なのは、西日本(沖縄を除く)の日本人です。西日本(沖縄を除く)の日本人→東日本の日本人→沖縄の日本人→アイヌ人の順にD系統が強くなっていきます。大陸から日本に農耕を伝えた人々の遺伝学的影響は、特に西日本(沖縄を除く)において顕著であるということです。
D系統はチベットと日本によく見られますが、チベットに見られるD系統と日本に見られるD系統は非常に遠い関係にあります。
前回の記事で、Y染色体DNAのC系統、D系統、F系統がアフリカ以外の世界に広がっていったことをお話ししました。しかし、アフリカの外の男性のY染色体DNAは、ほとんどがF系統です。F系統は、人数が多く、分布域が広いので、G、H、I、J、K・・・のような細かな下位系統に分類されています。それに対して、D系統は、人数が少なく、分布域が狭いので、そのように細かく分類されていません。
例えば、ヨーロッパによく見られるR系統とインディアンによく見られるQ系統の隔たりより、チベットによく見られるD系統と日本によく見られるD系統の隔たりのほうがずっと大きいです。ここでいう隔たりとは、分かれてから経過した時間のことです。
D系統は世界の中ですっかり稀少になっており、わずかな手がかりからその歴史を探らなければなりません。D系統の内部を細かく調べるための分析手法として、Short Tandem Repeatの話をします。Short Tandem Repeatは、生物学・人類学における重要な分析手法の一つです。
●Short Tandem Repeatを理解する
Short Tandem Repeat(ショートタンデムリピート)は、Microsatellite(マイクロサテライト)と呼ばれることもあります。
Y染色体DNAは、他のDNAと同じように、アデニンA、チミンT、グアニンG、シトシンCという四種類の物質が列を作っています。延々と続く長い列ですが、ところどころにある配列が繰り返されている箇所があります。
(注意! 以下では、Short Tandem Repeatのポイントをすばやく伝えるために、話を実際より単純にしてあります。)
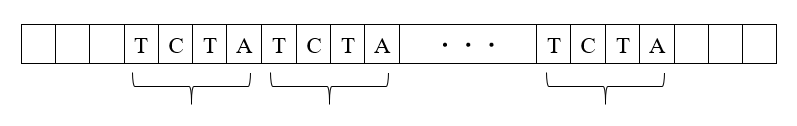
例えば、上の図ではTCTAという配列が10回繰り返されています。このような配列の繰り返しを、Short Tandem Repeatといいます。なぜShort Tandem Repeatが重要かというと、配列が繰り返されている箇所は、その他の箇所より変異(変化)しやすいのです。上の図ではTCTAという配列が10回繰り返されていますが、この10回という繰り返し回数が、遠くない将来に11回になったり、9回になったりします(繰り返し回数が1回増えたり1回減ったりするのが普通です)。さらに、11回だった繰り返し回数が12回になることもあるし、9回だった繰り返し回数が8回になることもあります。
上の図のTCTAという配列が繰り返されている領域を「繰り返し領域1」と呼ぶことにしましょう。同じようになんらかの配列が繰り返されている領域を次々に見つけ、それらに「繰り返し領域2、繰り返し領域3、繰り返し領域4、繰り返し領域5、繰り返し領域6、繰り返し領域7」と名前をつけます。繰り返し領域1ではTCTAという配列が何回か繰り返されている、繰り返し領域2では別の配列が何回か繰り返されている、繰り返し領域3ではさらに別の配列が何回か繰り返されている・・・という具合です。
繰り返し領域1で配列が10回繰り返されている
繰り返し領域2で配列が10回繰り返されている
繰り返し領域3で配列が10回繰り返されている
繰り返し領域4で配列が10回繰り返されている
繰り返し領域5で配列が10回繰り返されている
繰り返し領域6で配列が10回繰り返されている
繰り返し領域7で配列が10回繰り返されている
このようになっている場合に、それを「10‒10‒10‒10‒10‒10‒10」と書き表すことにします。D系統のY染色体DNAを持つ日本人男性10名を集め、繰り返し領域1~7を調べたところ、以下のようになっていたとしましょう。
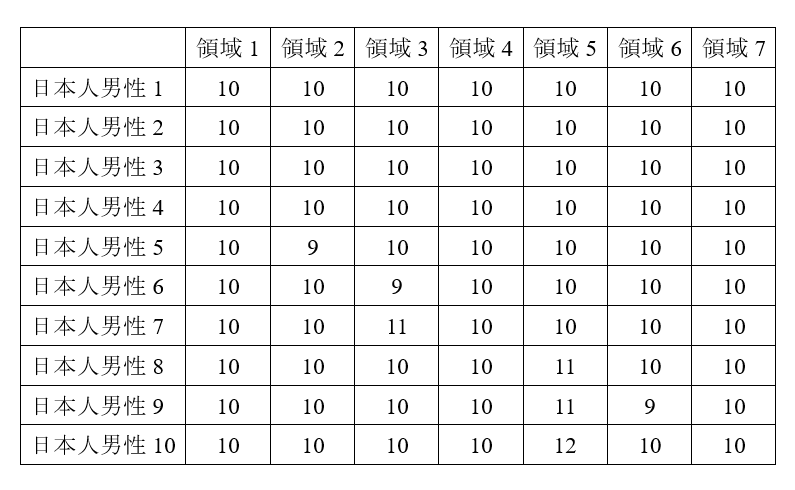
日本人男性1~4は「10‒10‒10‒10‒10‒10‒10」のタイプです。日本人男性5のタイプは、日本人男性1~4のタイプと比べて、領域2で数値が1減っています。日本人男性6のタイプは、日本人男性1~4のタイプと比べて、領域3で数値が1減っています。日本人男性7のタイプは、日本人男性1~4のタイプと比べて、領域3で数値が1増えています。日本人男性8のタイプは、日本人男性1~4のタイプと比べて、領域5で数値が1増えています。
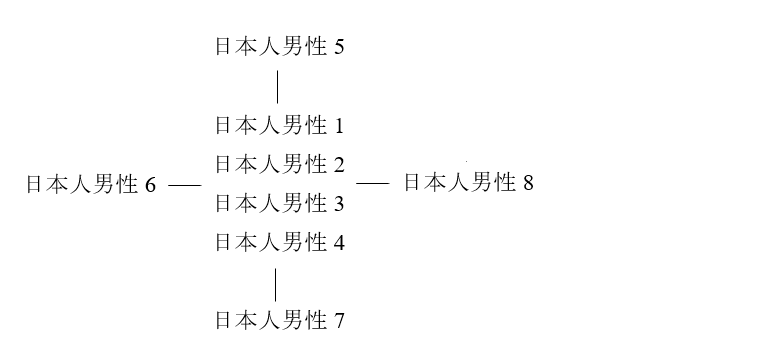
日本人男性5のタイプ、日本人男性6のタイプ、日本人男性7のタイプ、日本人男性8のタイプは、日本人男性1~4のタイプから1ステップずれていますが、ずれ方がそれぞれに異なります。日本人男性1~8は、以下のように書き表すことができます。
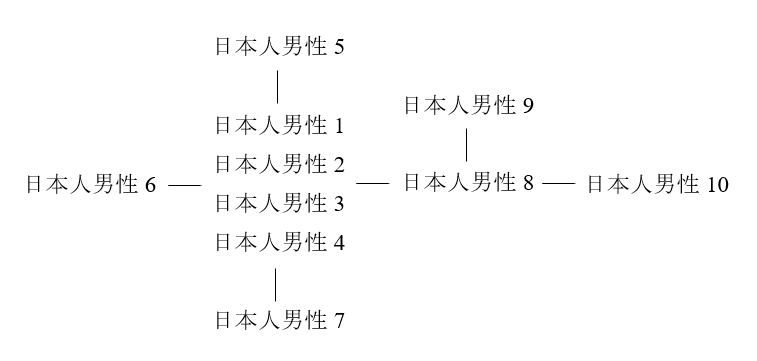
日本人男性9と日本人男性10はどうでしょうか。日本人男性9のタイプと日本人男性10のタイプは、日本人男性8のタイプから1ステップずれているタイプです。日本人男性9と日本人男性10は、以下のように書き表すことができます。
このように、Short Tandem Repeatを調べると、D系統のY染色体DNAを持つ日本人男性同士が互いにどのような関係にあるのか明らかにすることができます。Y染色体DNAの系統分類は繰り返し領域以外で行われますが、繰り返し領域も調べることによって細かな区別が可能になります。
大変興味深いことに、小金渕佳江氏らがD系統のY染色体DNAを持つアイヌ人と沖縄人のShort Tandem Repeatを調べています(図はKoganebuchi 2012より引用)。
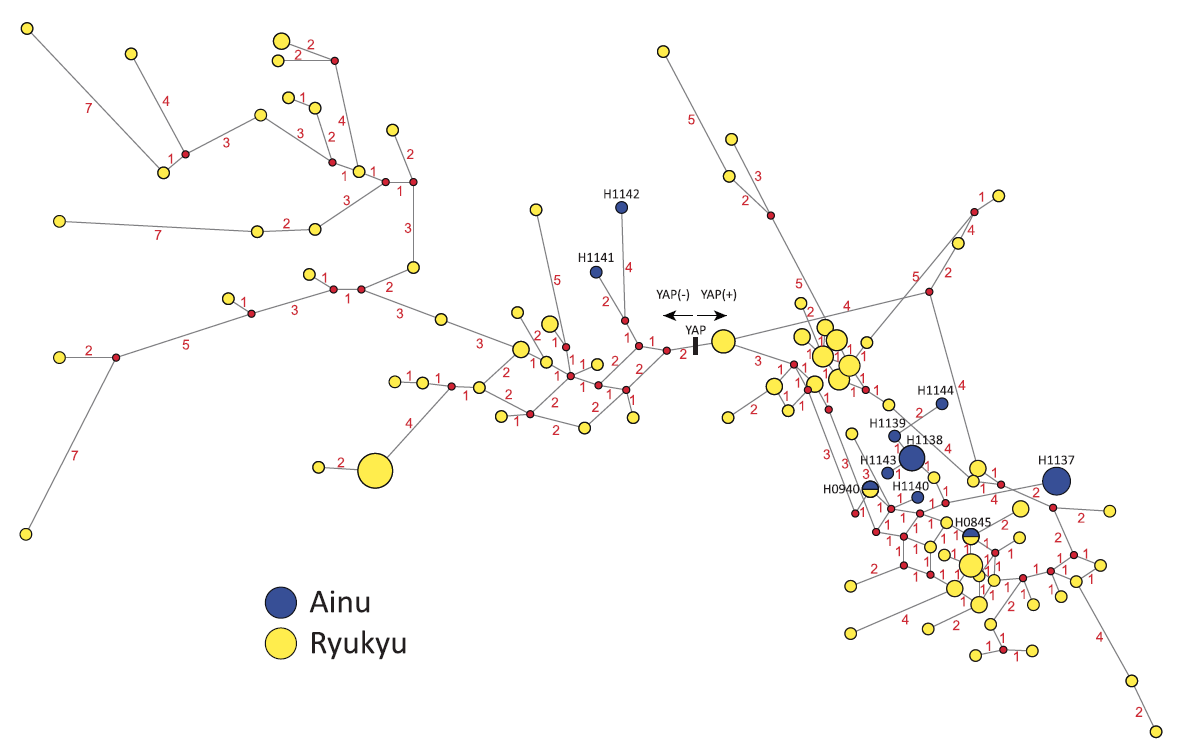
上に説明した要領で、アイヌ人男性と沖縄人男性のY染色体DNAのShort Tandem Repeatが調べられ、それらの男性が線で結ばれています。赤い数字は、何ステップ隔たっているかを示しています。YAP(+)と書いてある側が、Y染色体DNAのD系統のデータです。アイヌ人も沖縄人も含めて、日本に見られるD系統は、ほぼすべてD-M55という下位系統です。アイヌ人にD-M55系統が多く見られる、沖縄人にD-M55系統が多く見られるというところで話が終わってしまうことが多いですが、小金渕氏らの研究はさらに踏み込んでShort Tandem Repeatまで調べています。
見ての通り、Short Tandem Repeatのバリエーションはアイヌ人と沖縄人で大きく違います。沖縄人がアイヌ人に対してバリエーションの豊かさを見せています。沖縄人の調査人数に比べてアイヌ人の調査人数が少ないということもありますが、それを差し引いても、大きな差がありそうです。D系統は南から北へ広がっていった(つまり南にいた集団の一部が北に広がっていった)のではないかと思わせるデータです。
H. Shi氏らが東ユーラシアでD系統の男性のShort Tandem Repeatを広く調べていますが、チベットのD系統もとても北(中央アジアやモンゴル)から来たようには見えず、南から来たと考えざるをえません(Shi氏らが指摘しているように、Short Tandem Repeatに基づいて図を描いた時に、中央アジアやモンゴルのD系統の男性はことごとく、中心ではなく、末端に位置します。D系統に関しては、中央アジアやモンゴルからチベットに広がっているのではなく、チベットから中央アジアやモンゴルに広がっているのです)(Shi 2008)。
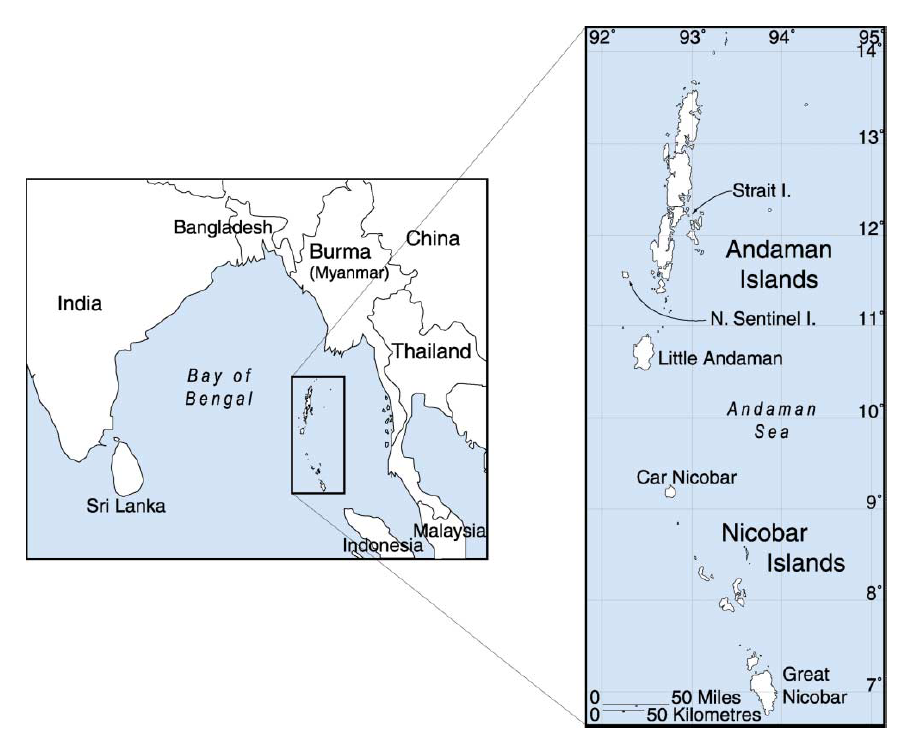
実は、チベットと日本のほかにもう一つ、D系統が多く見られる場所があります。それは、インド洋東部に浮かぶアンダマン諸島です(図はWikipediaより引用)。
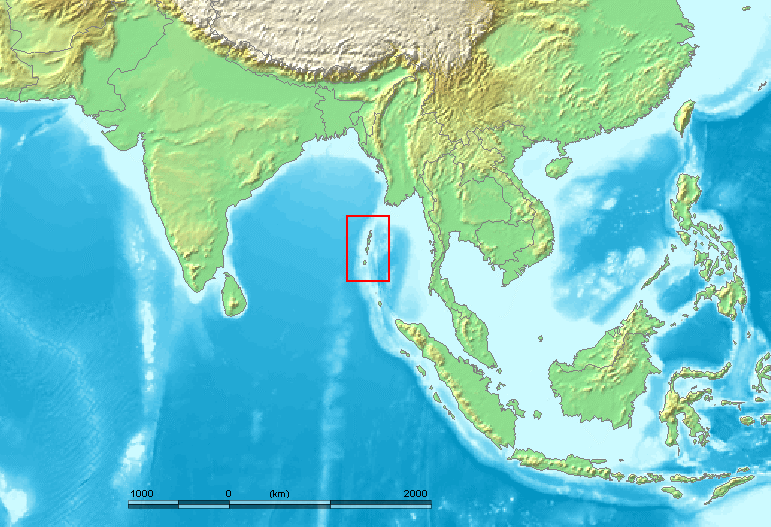
南から広がったように見える日本のD系統、南から広がったように見えるチベットのD系統、そしてアンダマン諸島に残るD系統・・・。こうなると、D系統はアフリカから中東に出て、中東から南アジアを通って東南アジアに達したのではないかと考えたくなります。これは、どこかで見たルートです。そうです、前回の記事でお話ししたC系統と同じルートです。しかし、不思議なことに、古くに東南アジアに達したC系統はオーストラリア・パプアニューギニア方面によく見られますが、同じく古くに東南アジアに達したはずのD系統はオーストラリア・パプアニューギニア方面に全然見られません。この問題はほとんど触れられないまま現在に至っていますが、東アジアの歴史を考えるうえで重要な問題と思われます。なぜそのようになっているのでしょうか。
補説
ついにアフリカで発見されたD系統
Y染色体DNAのD系統は東ユーラシアでしか見つかっていなかったため、D系統はアフリカで生じたのか、西ユーラシアで生じたのか、東ユーラシアで生じたのかという問題がありました。しかし、M. Haber氏らの研究によって、D系統のY染色体DNAを持つアフリカ(ナイジェリア)の男性がいることが明らかにされました(Haber 2019)。正確に言うと、これらの男性のY染色体DNAは、チベット、アンダマン諸島、日本などのD系統に極めて近いが、チベット、アンダマン諸島、日本などのD系統に共通しているM174という変異を持っていませんでした。M174という変異が起きる少し前に分かれて、そこからM-174という変異を経験しなかったのが今回のタイプ、M-174という変異を経験したのがチベット、アンダマン諸島、日本などのタイプということです。
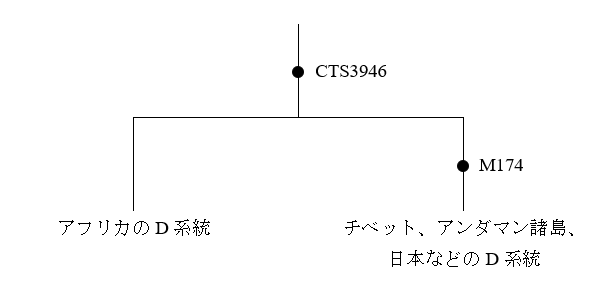
今回の発見により、D系統はM174という変異ではなく、CTS3946という変異によって定義されるようになりました。D系統と姉妹関係にあるE系統がアフリカで生じていることを考えると、上のCTS3946という変異はアフリカで起き、M174という変異はアフリカの外で起きた可能性が濃厚です。謎めくD系統の研究が一歩前進しました。
参考文献
Haber M. et al. 2019. A rare deep-rooting D0 African Y-chromosomal haplogroup and its implications for the expansion of modern humans out of Africa. Genetics 212(4): 1421-1428.
Hammer M. F. et al. 2006. Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes. Journal of Human Genetics 51(1): 47-58.
Koganebuchi K. et al. 2012. Autosomal and Y-chromosomal STR markers reveal a close relationship between Hokkaido Ainu and Ryukyu islanders. Anthropological Science 120(3): 199-208.
Nonaka I. et al. 2007. Y-chromosomal binary haplogroups in the Japanese population and their relationship to 16 Y-STR polymorphisms. Annals of Human Genetics 71(4): 480-495.
Shi H. et al. 2008. Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations. BMC Biology 6: 45.
Wang C. et al. 2013. Inferring human history in East Asia from Y chromosomes. Invetigative Genetics 4(1): 11.