

日本人に最も多く見られるY染色体DNAはO系統で、次に多く見られるのはD系統であると以前にお話ししました(アイヌ人と沖縄人のDNAを比べると・・・(Y染色体ハプログループDの研究)を参照、図はWang 2013より引用)。
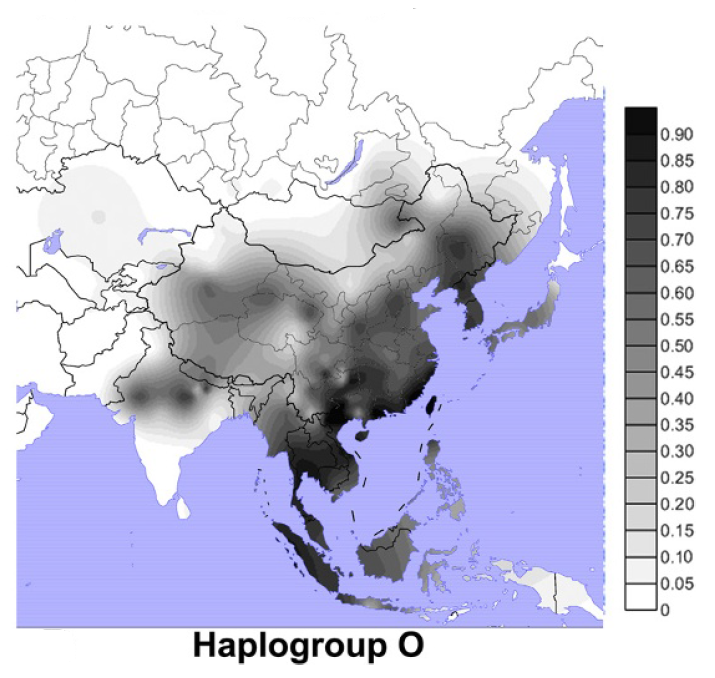
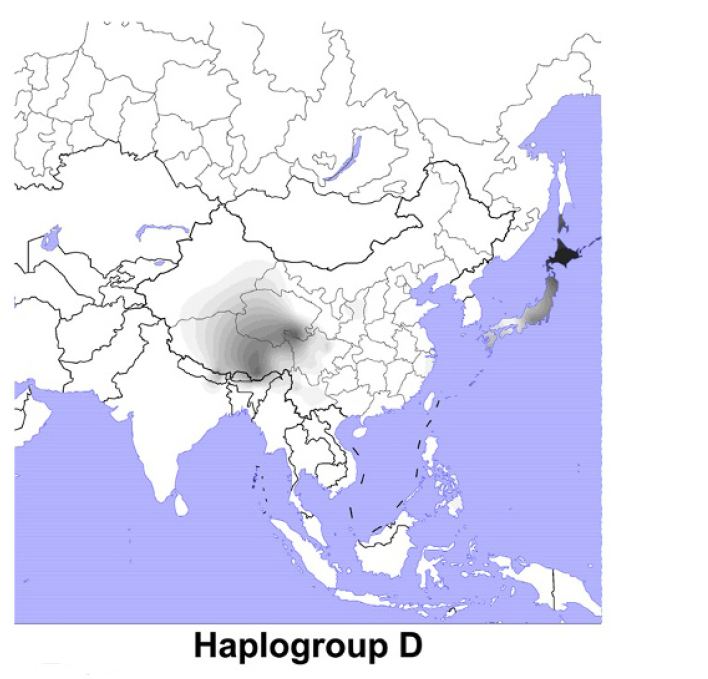
O系統は日本の近隣に非常に多く見られ、D系統は日本の近隣にほとんど見られません。この点で、O系統とD系統は対照的です。基本的に、O系統は弥生時代初めから稲作とともに日本列島に入ってきた系統で、D系統はそれ以前から日本列島に存在した系統と考えられます(縄文時代の途中からO系統が日本列島に入ってきていた可能性もありますが、話がそれてしまうので、その可能性はここでは脇に置いておきます)。
東アジア・東南アジアに見られるY染色体DNAの大部分はO系統で、O系統は大雑把に言うと以下の四つの下位系統に分けることができます(東アジア・東南アジア全体でごくわずかしか見られないマイナーな下位系統は図から省いてあります)。
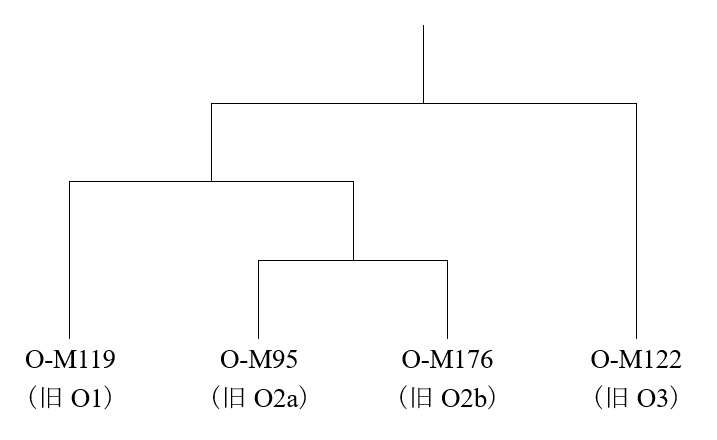
この中で、日本人に最も多く見られるのはO-M176(旧O2b)、次に多く見られるのはO-M122(旧O3)で、O-M119(旧O1)とO-M95(旧O2a)はわずかしか見られません(Nonaka 2007では、西日本(沖縄を除く)で36.1%、23.9%、4.1%、1%、関東で30.7%、14.5%、2.2%、0%となっており、沖縄人とアイヌ人を除けば、大体このようになりそうです)。日本人に最も多く見られるO-M176は特殊で、分布がほとんど朝鮮半島と日本列島に限られています(Xue 2006)。O-M176は、朝鮮半島と日本列島に多く見られ、稲作に関係がありそうなので、中国のどこかにも見られそうですが、中国には全くと言ってよいほど見られないのです。なぜでしょうか。
Li Hui氏らの興味深い研究があります(Li 2007)。長江文明が栄えていた頃の長江流域の住民のY染色体DNAを調べていますが、長江下流域ではO-M119が支配的で、長江中流域ではO-M95が支配的だったことが明らかになっています。これは興味深い結果です。稲作発祥の地である長江下流域と中流域で支配的だったO-M119とO-M95が、日本列島にはわずかしか見られないからです。
ちなみに、長江下流域で支配的だったO-M119(その主な下位系統としてO-M110があります)は、特に台湾(正確には台湾原住民)とフィリピンに多く見られます(Delfin 2011)。中国南部の福建省のあたりから台湾を経由してさらに南方に広がっていったオーストロネシア語族の話者と関係があると考えられます(台湾とオーストロネシア語族を参照)。
O-M119が日本にわずかしか見られないことから、長江下流域あるいはさらに南方にいた稲作民が日本に直接来ることがあったとしても、それが主流でなかったことは明らかです。
長江中流域で支配的だったO-M95は、現代では特にオーストロアジア語族(ベトナム系言語)の分布地域(インドシナ半島からインド内部にかけて)に多く見られ、マレーシアとインドネシア西部にもある程度見られます(Chaubey 2011)。
上に示したO系統の系統図を見ると、O-M95とO-M176が比較的近い関係にあるので、過去にはこんな考えもありました。長江中流域で稲作が始まり、そこにいた人間集団の一部がインドシナ半島に向かい、一部が朝鮮半島と日本列島に向かったのではないかという考えです。要するに、O-M95とO-M176の共通祖先型O-M268(旧O2)がかつて長江中流域に存在し、一下位系統であるO-M95がインドシナ半島に向かい、別の下位系統であるO-M176が朝鮮半島と日本列島に向かったのではないかと言いたいのです。しかし、長江中流域から稲作が広がった出来事と、Y染色体DNAのO-M268がO-M95とO-M176に分かれた出来事は、時間的に全然合いません。長江中流域から稲作が広がったのは、せいぜい過去1万年ぐらいのことですが、Y染色体DNAのO-M268がO-M95とO-M176に分かれたのは、2.5~3万年前ぐらいのことです(www.23mofang.com/ancestry/ytree/O-M268を参照、O-M176はO-P49と記されることもあります)。稲作開始よりはるか前に、O-M95とO-M176は分かれていたのです。稲作開始の時点では、O-M95を持つ人間集団が長江中流域で稲作を行い、O-M176を持つ人間集団はいくらか離れたところにいて、稲作を行っていなかったと見られます。O-M268の下位系統は長江下流域に見られないので、O-M268が中国南部の内陸に存在し、そこからインドシナ半島方面と朝鮮半島・日本列島方面に分かれたのではないかというG. Chaubey氏らの考えは、的を射ているのではないかと思われます(Chaubey 2020)。
本記事で示したのは、一例にすぎません。そもそも、人と稲作と言語の広がりは一致するとは限らないのです(稲作のところをその他の文明・文化的特徴に置き換えても、同じことが言えます)。確かな根拠がないのに、人と稲作と言語の広がりは常に一体だと思い込んでしまうから、日本人の起源探しも、稲作の起源探しも、日本語の起源探しも難航するのです。日本の例でも明らかになってきましたが、なぜ人と稲作と言語の広がりが必ずしも一致しないのか考えてみることにしましょう。
ほぼ朝鮮半島と日本列島に限られているY染色体DNAのO-M176がどこから来たのかという問題も、興味深い問題であり、中核的な問題です。
参考文献
Chaubey G. et al. 2011. Population genetic structure in Indian Austroasiatic speakers: the role of landscape barriers and sex-specific admixture. Molecular Biology and Evolution 28(2): 1013-1024.
Chaubey G. et al. 2020. Munda languages are father tongues, but Japanese and Korean are not. Evolutionary Human Sciences 2: e19.
Delfin F. et al. 2011. The Y-chromosome landscape of the Philippines: Extensive heterogeneity and varying genetic affinities of Negrito and non-Negrito groups. European Journal of Human Genetics 19(2): 224-230.
Li H. et al. 2007. Y chromosomes of prehistoric people along the Yangtze River. Human Genetics 122(3-4): 383–388.
Nonaka I. et al. 2007. Y-chromosomal binary haplogroups in the Japanese population and their relationship to 16 Y-STR polymorphisms. Annals of Human Genetics 71(4): 480-495.
Wang C. et al. 2013. Inferring human history in East Asia from Y chromosomes. Invetigative Genetics 4(1): 11.
Xue Y. et al. 2006. Male demography in East Asia: A north-south contrast in human population expansion times. Genetics 172(4): 2431-2439.