
以前に、古代北ユーラシアの人々のDNAについてお話ししたことがありました(一貫性を示す古代北ユーラシアの人々のDNAを参照)。その時はミトコンドリアDNAとY染色体DNAの話しかしませんでしたが、さらにDNA全体が調べられ、古代の様子が一層明らかになってきました(図はMassilani 2020より引用)。
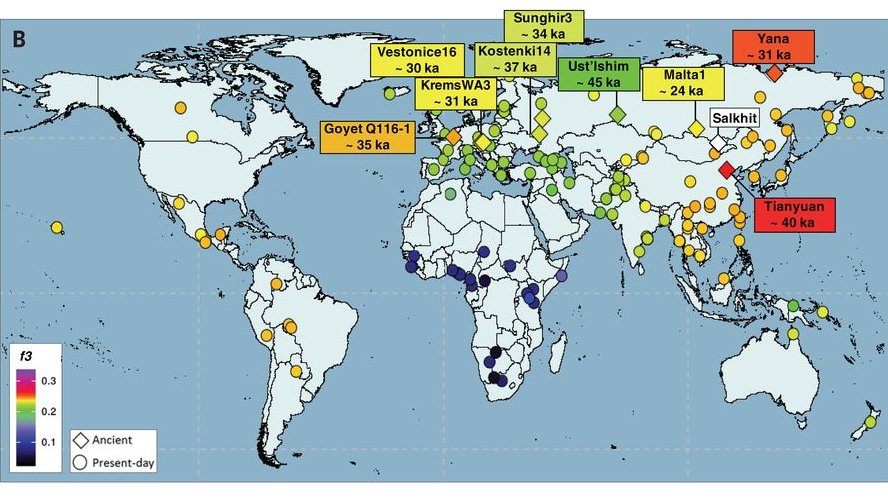
今のところ、東アジアの歴史を考えるうえで特に重要なのは、以下の6名です。
Ust’-Ishim 45000年前頃のロシアのウスチイシムの男性
Tianyuan 40000年前頃の中国の田園洞遺跡の男性
Salkhit 34000年前頃のモンゴルのサルヒトの女性
Yana 31000年前頃のロシアのやヤナ遺跡の男性2名
Mal’ta 24000年前頃のロシアのマリタの男性
Ust’-Ishimの男性、Yanaの男性2名、Mal’taの男性についてはすでに、ミトコンドリアDNAとY染色体DNAのデータに基づいて、中東から中央アジアに向かい、中央アジアから北ユーラシアに拡散していった人々だろうと述べました。DNA全体を調べたMassilani氏らの研究(Massilani 2020)でも、そのことが確かめられました。対照的に、Tianyuanの男性とSalkhitの女性は、中東から東南アジアに向かい、東南アジアから北上してきた人々であることが明らかになりました。
Salkhitの女性のDNAの75%は東南アジアルート(南方ルート)から来ており、25%は中央アジアルート(北方ルート)から来ていると分析されています(Massilani 2020)。やはり、東南アジアルートを通ってやって来た人々と中央アジアルートを通ってやって来た人々は、非常に古くから混ざり合っていたのです。東アジアの歴史の初期の頃から、そのような混ざり合いが起きていたのです。これはとても重要なことです。これで考古学的証拠と生物学的証拠が完全に嚙み合いました(考古学的証拠については、4万年前の東アジアを参照)。
Yanaの男性2名は31000年前頃にユーラシア大陸の最北部にいた人たちで、ともにミトコンドリアDNAがU系統、Y染色体DNAがP系統であることがわかっていました(Sikora 2019)。このミトコンドリアDNAのU系統とY染色体DNAのP系統は、中東→中央アジア→北ユーラシアと進出したものです。しかし、男性2名のDNA全体を調べると、その30%以上が東南アジアルートから来ています(Massilani 2020)。ミトコンドリアDNAとY染色体DNAは重要ですが、それだけでは見えない部分があり、その見えない部分がかなり大きい場合もあることをはっきりと示しています。
日本人の起源について論じる時や日本語の起源について論じる時に、よく「北方」と「南方」という言葉が使われてきました。しかし、筆者は思っていました。その「北方」とはなんなのか、「南方」とはなんなのかと。よくわかっていない「北方」と「南方」によって、日本人の起源や日本語の起源を明らかにできるのかと。
「北方」と「南方」が曖昧模糊としているのも問題ですが、もう一つ問題なのは、旧石器時代への関心が低いことです。弥生時代と縄文時代に比べて、旧石器時代はなかなか注目されません。確かに、旧石器時代は手がかりが少ないので、無理もないかもしれません。
日本は、旧石器時代→縄文時代→弥生時代と変遷しました。しかし、旧石器時代から縄文時代への変化と、縄文時代から弥生時代への変化は、かなり異質なようです。縄文時代から弥生時代への変化では、(水田)稲作が行われるようになったり、金属器(青銅器、鉄器)が使われるようになったりしました。これは、大陸からやって来た人々が引き起こした変化です。旧石器時代から縄文時代への変化は、どうでしょうか。旧石器時代から縄文時代への変化では、土器が作られるようになったり、定住が始まったりしました。
東アジアでは農耕が始まるずっと前から土器が作られており、どこで土器が作られ始めたのか盛んに研究されてきました。考古学調査が進み、年代測定技術が発達するにつれて、不思議な様相が浮かび上がってきました(図はKuzmin 2015より引用)。
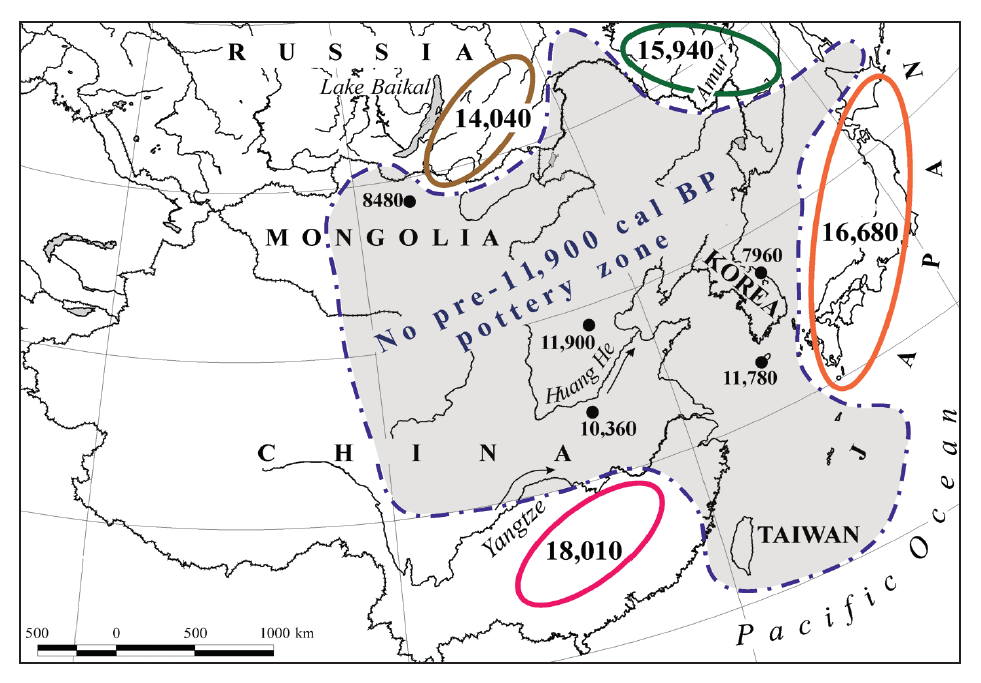
14000~18000年前頃から、中国南部、日本、アムール川流域、バイカル湖周辺に土器が現れます(中国南部にもっと古い報告例がありますが、広く認められていません(Wu 2012、Kuzmin 2013))。しかし、これらの地域の間には同時代の土器が全然見られず、土器が一つの地域で生まれて残りの地域に伝わったという説明は困難な状況です。朝鮮半島にいたっては、8000年前頃になるまで土器が現れません。
日本で最古の土器は、青森県の大平山元遺跡(おおだいやまもといせき)で見つかっていますが、これも、他の地域から伝わってきたという説明は困難です。日本最古の土器は、縄文(縄を押しつけてつける模様)がなく、無文です。
一般に、ある地域に新しい現象または状況が見られ始めた場合、以下のパターンが考えられます。
(1)よそから大勢の人々が入って来て、変化が生じた。
(2)よそから少数の人々が入って来て、変化が生じた。
(3)よそから人は入って来ないで、純粋にその地域の内部で変化が生じた。
現状では、日本の土器作りは(3)と考えるしかありません。日本の場合には、土器よりいくらか遅れて定住が始まりました。日本最古の土器は東北で見つかっていますが、定住は九州から広がっていきました(Pearson 2006)。土器と定住は縄文時代の重要な特徴ですが、これらはいっしょに発生していっしょに広がっていったわけではないようです。いずれにせよ、日本の旧石器時代から縄文時代への変化は、よそから人々が全く入って来ないあるいはほとんど入って来ない変化であったと見られます。そうなると、縄文時代の人々については考えるが旧石器時代の人々については考えないというのは、なんともおかしなことです。連続しているわけですから。
ただ、上の地図が示しているように、これほど近い時期に東アジアの四つの地域で土器が現れたのはなぜかという疑問は残ります。単なる偶然として片づけられるようなものでもないと思われます。Kuzmin氏などは土器が生まれたのは食材の調理と保存のためではないかと述べていますが、その可能性は高いでしょう(Kuzmin 2013)。実際、日本の縄文時代草創期(縄文時代の最も古い時期)の土器の付着物が調べられ、土器が特に淡水・海洋生物の調理に使われていたことが明らかになっています(Craig 2013)。
筆者は、マンモスのような大型の動物が獲れなくなってきて、人々の注意・関心が陸上のその他の動物、植物、魚介類などの様々な対象に強く向かうようになったことが大きかったのではないかと考えています。様々な対象の中には、そのまま食べるのに不向きなものも多かったでしょう。東アジアの主食になったアワとイネにしたって、そのまま食べるのに向いていません。土器の始まり、定住の始まり、農耕の始まりについては膨大な議論がありますが、いずれも究極的には下線部の事情と無関係でないと思われます(ちなみに、日本では土器、定住、農耕の順に見られましたが、必ずその順で見られるとは限りません(Gibbs 2016))。
日本人の起源や日本語の起源を知りたければ、日本とその周辺地域を含む旧石器時代からきちんと考えなければなりません。筆者がこのブログでやっていることを見ればわかると思いますが、日本語の起源というのは、日本語はどこから来たのかというより、日本語の一語一語はどこから来たのかという問題です。日本語の一語一語がどこから来たのか明らかにするためには、人類の歴史がどのように展開してきたのかよく知る必要があります。
参考文献
Craig O. E. et al. 2013. Earliest evidence for the use of pottery. Nature 496(7445): 351-354.
Gibbs K. et al. 2016. A comparative perspective on the ‘western’ and ‘eastern’ Neolithics of Eurasia: Ceramics; agriculture and sedentism. Quaternary International 419: 27-35.
Kuzmin Y. V. 2013. Origin of Old World pottery as viewed from the early 2010s: When, where and why? World Archaeology 45(4): 539-556.
Kuzmin Y. V. 2015. The origins of pottery in East Asia: Updated analysis (the 2015 state-of-the-art). Documenta Praehistorica 42: 1-11.
Massilani D. et al. 2020. Denisovan ancestry and population history of early East Asians. Science 370(6516): 579-583.
Pearson R. 2006. Jomon hot spot: Increasing sedentism in south-western Japan in the Incipient Jomon (14,000–9250 cal. BC) and Earliest Jomon (9250–5300 cal. BC) periods. World Archaeology 38(2): 239-258.
Sikora M. et al. 2019. The population history of northeastern Siberia since the Pleistocene. Nature 570(7760): 182-188.
Wu X. et al. 2012. Early pottery at 20,000 years ago in Xianrendong Cave, China. Science 336(6089): 1696-1700.