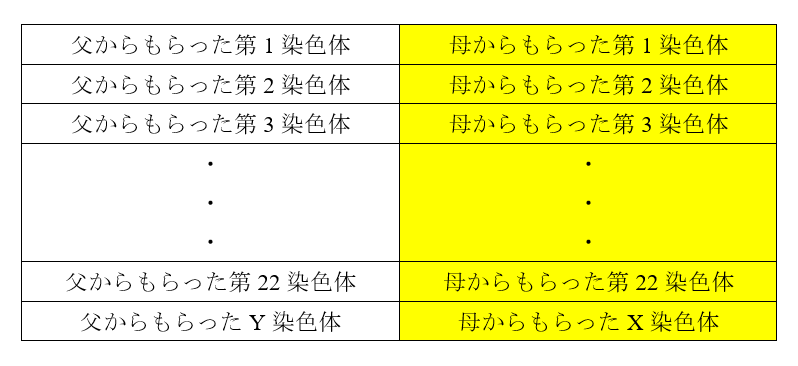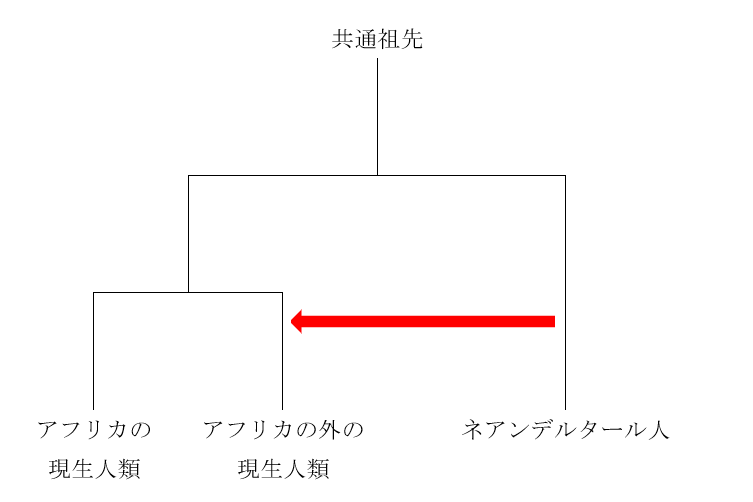

現生人類のDNAには、ネアンデルタール人由来の部分と、デニソワ人由来の部分が見られます。しかし、ネアンデルタール人由来の部分はアフリカの外の人々に広く見られますが、デニソワ人由来の部分はもっと分布が限られています。デニソワ人由来の部分が見られるのは、東ユーラシア、オセアニア、南北アメリカの人々です。要するに、アフリカを出て、東にどんどん進んでいった人々に見られるのです。
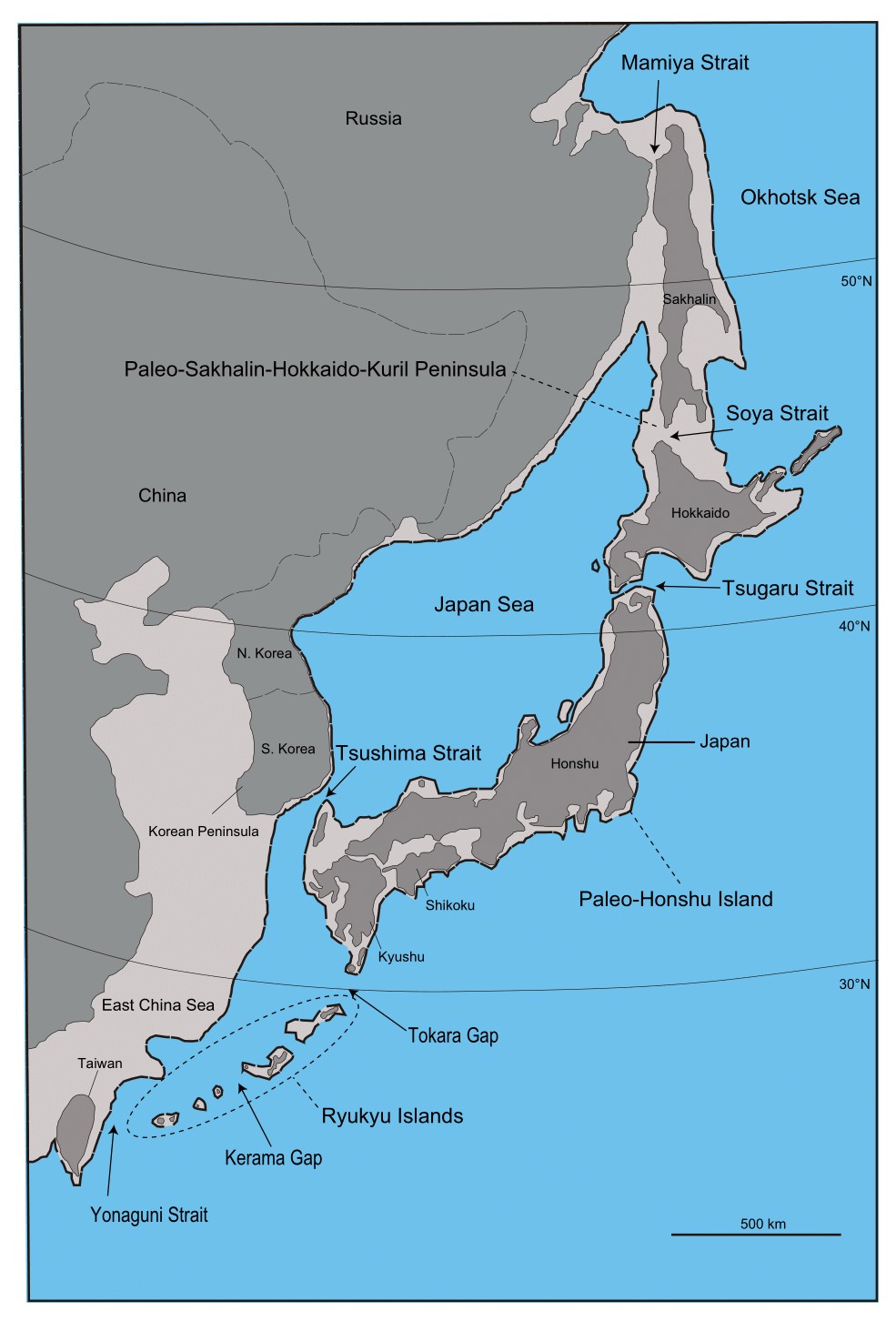
ロシアのアルタイ地方のデニソワ洞窟でデニソワ人が発見された後、デニソワ人のDNAが調べられました。そして、デニソワ人のDNAが現生人類に入っているかどうかが注目されました。ここで真っ先に浮かび上がってきたのが、なんとパプアニューギニアとオーストラリア(アボリジニ)の人々でした(Reich 2010、2011)。しかし、パプアニューギニアとオーストラリアの人々にデニソワ人のDNAが顕著に認められるというのは、ちょっと不可解な話でした。デニソワ人が発見されたデニソワ洞窟はシベリアにあるからです(地図はThe Guardian様のウェブサイトより引用)。
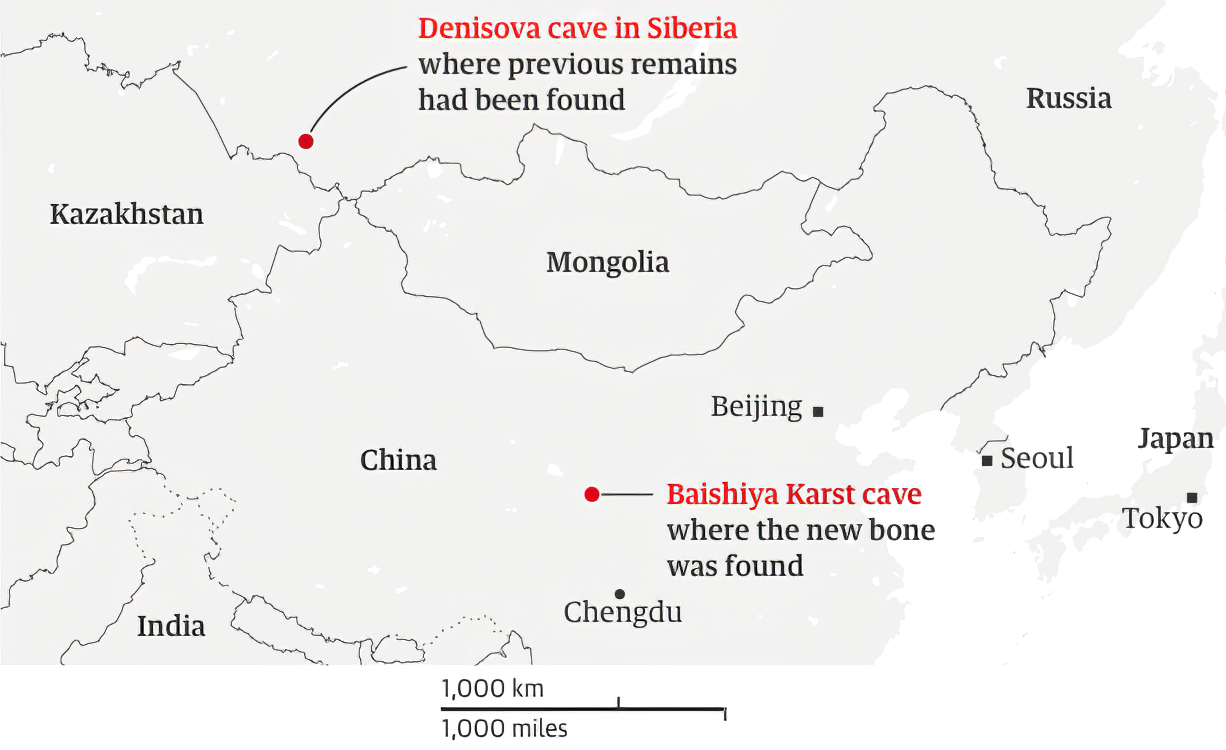
※Denisova Cave(デニソワ洞窟)は、カザフスタン、ロシア、モンゴルが集まっているあたりにあります。Baishiya Karst Cave(白石崖鍾乳洞)については後述します。
その後の研究によって、パプアニューギニアとオーストラリアの人々ほどではないが、東ユーラシア(東南アジア、東アジア、シベリア)と南北アメリカ(インディアン)の人々にも、デニソワ人のDNAがわずかに認められることがわかりました(デニソワ人のDNAは、パプアニューギニアとオーストラリアの人々では5%程度、東ユーラシアと南北アメリカの人々では0.2%程度です)(Prüfer 2014、Qin 2015)。
どうやら、デニソワ人は広い範囲に分布していたようです。デニソワ人が広い範囲に分布していたのであれば、シベリアでデニソワ人の骨が見つかったことと、パプアニューギニアとオーストラリアの人々のDNAにデニソワ人由来の部分が顕著に認められることは、矛盾しません。デニソワ人は北のほうにもいたし、南のほうにもいたということです。
しかしながら、デニソワ人が広く分布しているところに現生人類が流れ込んだとなると、複雑な展開が予想されます。デニソワ人と現生人類の接触は複数の地域で起きたかもしれません。そのような可能性がS. R. Browning氏らによって提起されました(Browning 2018)。Browning氏らは、パプアニューギニアの人々のDNAに見られるデニソワ人由来の部分と、東アジアの人々(中国人および日本人)のDNAに見られるデニソワ人由来の部分を比較し、以下のような見解を示しています。
・パプアニューギニアの人々のDNAに見られるデニソワ人由来の部分は、ロシアのアルタイ地方のデニソワ洞窟で見つかったデニソワ人と遠い関係にあるデニソワ人集団から入ったものである。
・東アジアの人々のDNAに見られるデニソワ人由来の部分は、ロシアのアルタイ地方のデニソワ洞窟で見つかったデニソワ人と遠い関係にあるデニソワ人集団から入ったものと、ロシアのアルタイ地方のデニソワ洞窟で見つかったデニソワ人と近い関係にあるデニソワ人集団から入ったものが混在している。
G. S. Jacob氏らも似た見解を示していますが、もっと複雑です(Jacob 2019)。Browning氏らは、「パプアニューギニアの人々と東アジアの人々にDNAを与えたデニソワ人集団」と「東アジアの人々だけにDNAを与えたデニソワ人集団」という二種類のデニソワ人集団によって説明しようとしていますが、Jacob氏らは、「パプアニューギニアの人々と東アジアの人々にDNAを与えたデニソワ人集団」と「東アジアの人々だけにDNAを与えたデニソワ人集団」と「パプアニューギニアの人々だけにDNAを与えたデニソワ人集団」という三種類のデニソワ人集団によって説明しようとしています。
Jacob氏らは、各地に存在したデニソワ人集団は35万年前頃から枝分かれし始めていたと推定しています(Jacob 2019)。互いに大きく隔たった複数のデニソワ人集団が、現生人類にDNAを与えたことは間違いないようです。
2008年にロシアのアルタイ地方のデニソワ洞窟で最初のデニソワ人が発見され、その後もデニソワ洞窟でデニソワ人の発見が相次ぎました(Reich 2010、Sawyer 2015、Slon 2017b)。そしてついに、デニソワ洞窟以外の場所でデニソワ人が発見されました(Chen 2019)。その発見場所が、冒頭の地図に示されたBaishiya Karst Cave(白石崖鍾乳洞、はくせきがいしょうにゅうどう)です。
白石崖鍾乳洞は、チベット高原の一番北東にあります。ヒマラヤ山脈から遠く離れていますが、それでも標高3280 mにあります。ちなみに、デニソワ洞窟は標高700 mにあります。白石崖鍾乳洞でのデニソワ人の発見は、大きな驚きをもたらしました。見つかったのは歯の付いた下あごの骨ですが、これが16万年以上前のものと推定されたのです(Chen 2019)。チベット高原は、気温が低いだけでなく、空気が薄く、かなり過酷なところです(チベット高原の平均標高は4000 mぐらいで、この高さでは酸素の量が低地の2/3未満になっています。酸素の量が1/3になると、エベレストの頂上並みで、人間はとても生活できません)。チベット高原には3~4万年頃から現生人類が現れましたが、それまではだれもいなかったと考えられていました(Zhang X. 2018)。そのチベット高原から、16万年以上前のデニソワ人の骨と歯が出たのです。Chen氏らの研究では、DNA分析がうまくいかず、タンパク質分析によって骨と歯がデニソワ人のものであると判定されました(Chen 2019、DNAとタンパク質の関係については補説を参照)。
時にDNA分析の代わりになりうるタンパク質分析は大きな進歩ですが、最近ではもっとびっくりする研究も出てきています。V. Slon氏らの研究などがそうです(Slon 2017a)。これまでは、古代人の骨と歯が見つかって、その骨と歯が調べられてきました。しかし、そのような発見はまれで、なかなか歴史を明らかにできないもどかしい状況がありました。Slon氏らの研究では、骨と歯が見当たらない場所でも、そこの堆積物から古代の人と動物のDNAを明らかにしてしまいます(人と動物の残骸が微細な形で残っているということです)。なんとも奇抜な研究ですが、このような研究が広まりつつあります。
最近の研究の進歩は、日本の研究にとっても朗報かもしれません。日本は火山が非常に多く(富士山もその一つです)、それらの火山は長い歴史の中で何回も何回も噴火してきました。火山から噴出して降ってくる火山灰のために、日本の土壌は酸性度が高く、骨と歯がなかなか残りません。沖縄を除く本土は特にそうです。
Chen氏とSlon氏らは、すでに白石崖鍾乳洞でも、堆積物から何人かのデニソワ人のDNAを明らかにしています(Zhang D. 2020)。年代の違う各層からデニソワ人のDNAが出ており、デニソワ人が何万年もの長期にわたって白石崖鍾乳洞にいたことが示されています。DNA分析により、白石崖鍾乳洞にいたデニソワ人がデニソワ洞窟で最初に見つかったデニソワ人と近い関係にあることも示されています。
冒頭の地図をもう一度見てください。デニソワ人が何万年もの長期にわたってデニソワ洞窟にいたことがわかっています(Douka 2019)。デニソワ人が何万年もの長期にわたって白石崖鍾乳洞にいたこともわかりました。デニソワ人がこの二箇所だけにいたとは到底考えられません。調査・分析技術が進歩しているので、今後東ユーラシアの様々な場所でデニソワ人の存在が確認される可能性が高いです。Browning氏らやJacob氏らは、パプアニューギニアの人々のDNAには見られないが東アジアの人々のDNAには見られるデニソワ人由来の部分があることを示していますが、このデニソワ人由来の部分がどこで東アジアの人々に入ったのかというのも気になります。デニソワ洞窟のあるシベリアでしょうか、それとも、中国南部でしょうか(Chen氏らは、白石崖鍾乳洞のデニソワ人の骨が、台湾近海で網にかかって発見された古人類の骨とよく似ているとも述べています(Chen 2019およびそこに挙げられているChang 2015))。
現生人類が東南アジアルート(南方ルート)と中央アジアルート(北方ルート)(それぞれが単純な一本のルートであるとは限りません)を通って東アジアに入ってくる4~5万年前頃は、東アジアの歴史における最重要局面と言っても過言ではありません。そのような重要な時代ではありますが、手がかりの少ない時代でもあります。現生人類がデニソワ人と接し、交わっていた時代でもあり、デニソワ人が貴重な情報を与えてくれるかもしれません。
現生人類が複雑な様相を呈しながら東アジアに入ってくる旧石器時代のことを無視して、日本の縄文時代および弥生時代の人々の素性を明らかにすることはできないのです。
補説
DNAとタンパク質の関係
見つかった骨または歯が、現生人類のものなのか、ネアンデルタール人のものなのか、デニソワ人のものなのか、それともその他の種のものなのか知りたければ、DNA配列を調べるのが一番です。しかし、それがかなわない時には、タンパク質を調べる方法もあります。DNA配列とタンパク質には密接な関係があるからです。
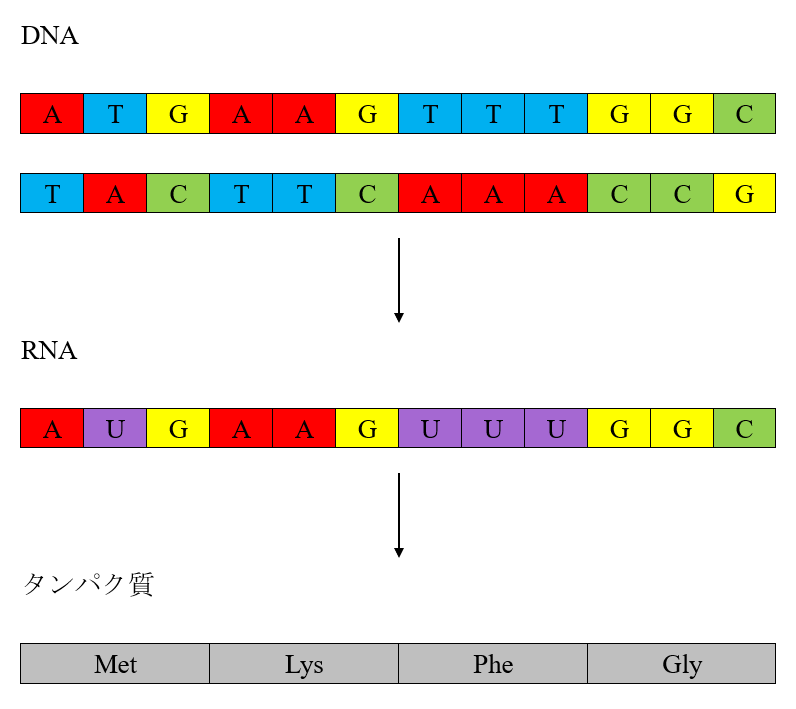
DNA配列に基づいてRNA配列が作られ、RNA配列に基づいてアミノ酸の連なりが作られ、このアミノ酸の連なりがタンパク質であるという関係があります。
それほど難しい話ではありません。図中のDNA配列を見てください。DNA配列は、アデニンA、チミンT、グアニンG、シトシンCという四種類の物質が作る列です。図のように、二つの列が寄り添うように存在しています。Aの向かいはT、Tの向かいはA、Gの向かいはC、Cの向かいはGと決まっています。二つの列のうちの一つの列(ここでは下段の列)をもとにして、RNA配列が作られます。
下段の列をもとにしてRNA配列が作られる時には、Aの向かいはU、Tの向かいはA、Gの向かいはC、Cの向かいはGになります。こうして、RNA配列は、アデニンA、ウラシルU、グアニンG、シトシンCという四種類の物質が作る列になります。RNA配列のAUGの部分が、Met(メチオニン)というアミノ酸を作りなさいという命令になっています。同様に、AAGの部分がLys(リジン)というアミノ酸を作りなさいという命令、UUUの部分がPhe(フェニルアラニン)というアミノ酸を作りなさいという命令、GGCの部分がGly(グリシン)というアミノ酸を作りなさいという命令になっています。このようにしてアミノ酸が多数連なったのが、タンパク質です(図には書ききれませんでしたが、タンパク質を構成するアミノ酸は20種類あります。タンパク質は、折りたたまれて立体構造を取っているのが普通です)。
一連の過程を見ればわかるように、おおもとのDNA配列が、どのようなアミノ酸の連なりを作り出すか、つまり、どのようなタンパク質を作り出すかを決定しています。現生人類、ネアンデルタール人、デニソワ人およびその他の種のDNA配列の違いは、作り出されるタンパク質の違いとして表れます。骨または歯からDNAが抽出できない時に、タンパク質を抽出して種の判別が行えるのは、そのためです。
参考文献
Browning S. R. et al. 2018. Analysis of human sequence data reveals two pulses of archaic Denisovan admixture. Cell 173(1): 53-91.
Chang C. et al. 2015. The first archaic Homo from Taiwan. Nature Communications 6(1): 6037.
Chen F. et al. 2019. A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau. Nature 569(7756): 409-412.
Douka K. et al. 2019. Age estimates for hominin fossils and the onset of the Upper Palaeolithic at Denisova Cave. Nature 565(7741): 640-644.
Jacob G. S. et al. 2019. Multiple deeply divergent Denisovan ancestries in Papuans. Cell 177(4): 1010-1021.
Prüfer K. et al. 2014. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 505(7481): 43-49.
Qin P. et al. 2015. Denisovan ancestry in East Eurasian and Native American populations. Molecular Biology and Evolution 32(10): 2665-2674.
Reich D. et al. 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468(7327): 1053-1060.
Reich D. et al. 2011. Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania. American Journal of Human Genetics 89(4): 516-528.
Sawyer S. et al. 2015. Nuclear and mitochondrial DNA sequences from two Denisovan individuals. Proceedings of the National Academy of Sciences of the United States of America 112(51): 15696-15700.
Slon V. et al. 2017a. Neandertal and Denisovan DNA from Pleistocene sediments. Science 356(6338): 605-608.
Slon V. et al. 2017b. A fourth Denisovan individual. Science Advances 3(7): e1700186.
Zhang D. et al. 2020. Denisovan DNA in Late Pleistocene sediments from Baishiya Karst Cave on the Tibetan Plateau. Science 370(6516): 584-587.
Zhang X. et al. 2018. The earliest human occupation of the high-altitude Tibetan Plateau 40 thousand to 30 thousand years ago. Science 362(6418): 1049-1051.