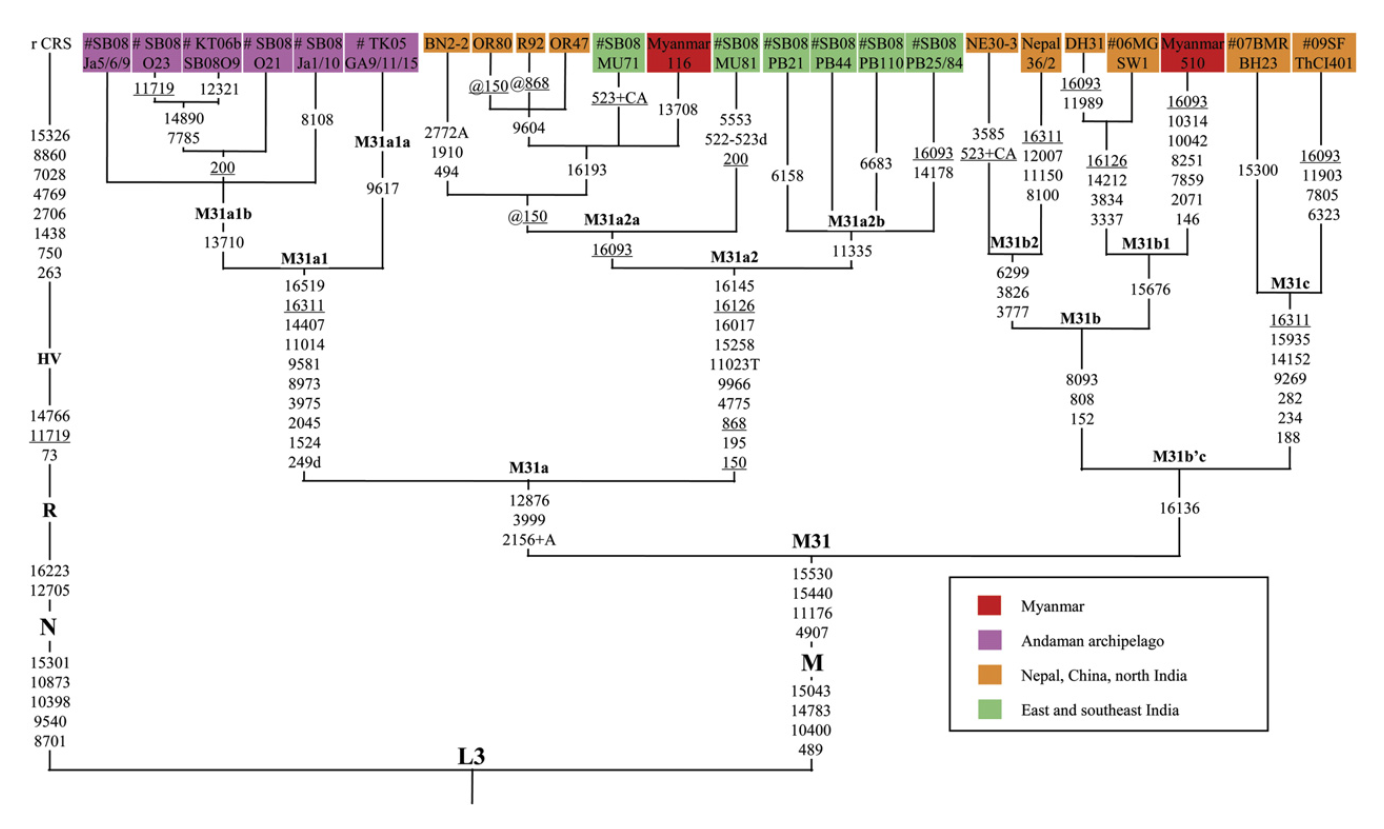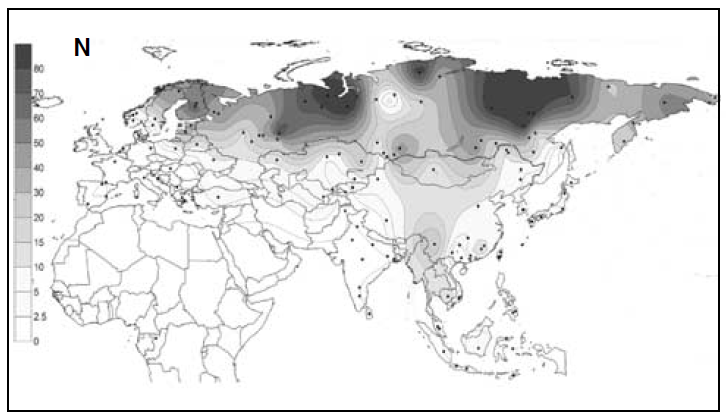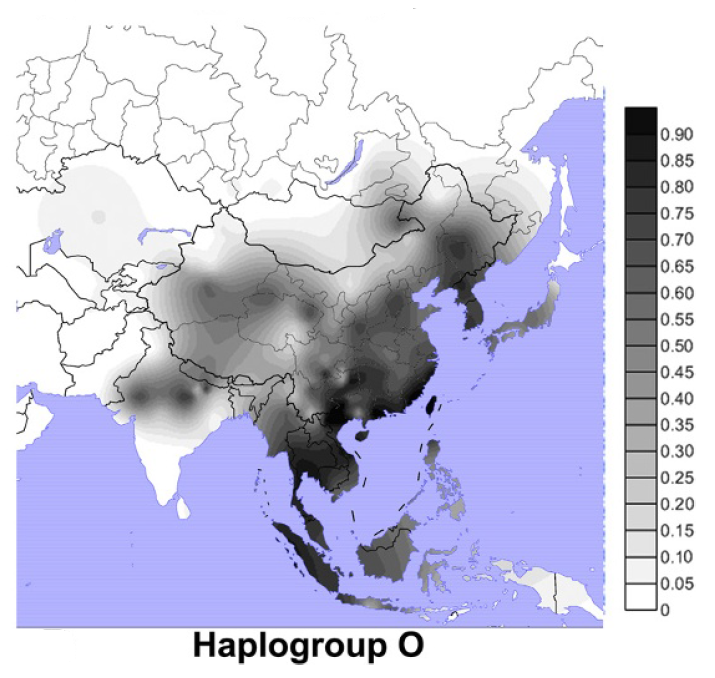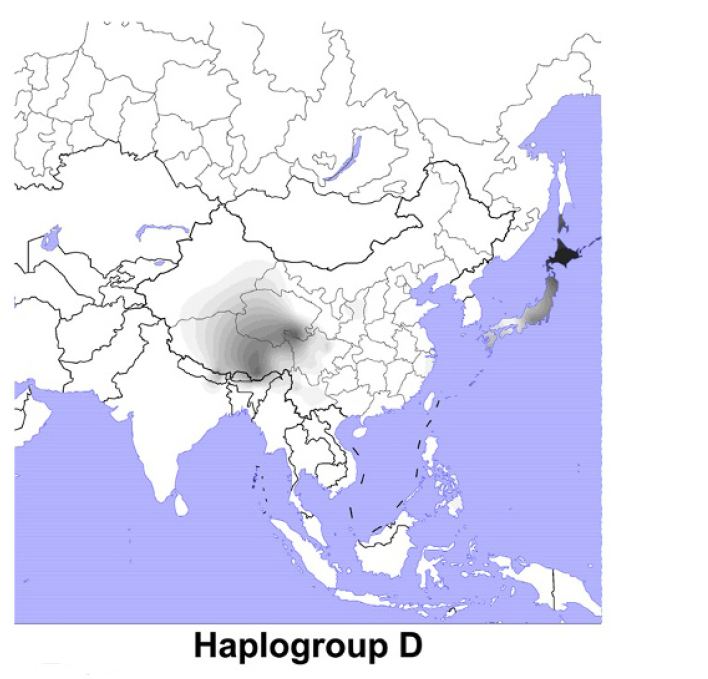

以前に古代北ユーラシアの人々のDNAの話をしましたが(一貫性を示す古代北ユーラシアの人々のDNAを参照)、ミトコンドリアDNAとY染色体DNAだけでなく、DNA全体の研究も進んでおり、新たな発見が続々と発表されています。いくつかの貴重な研究を紹介しますが、まずその前にDNAに関する基本事項を確認しておきましょう。
●ミトコンドリアDNAとY染色体DNAはDNA全体のごく一部にすぎない
アデニンA、チミンT、グアニンG、シトシンCという四種類の物質が長い長い列を作っています。このA、T、G、Cが作る暗号のような列が、私たちが持っている遺伝情報です。A、T、G、Cが作る暗号のような列は、どこにあるのでしょうか。皆さんも生物学の授業などで以下のような図を見たことがあるかと思います(図はパンタグラフ様のウェブサイトより引用)。
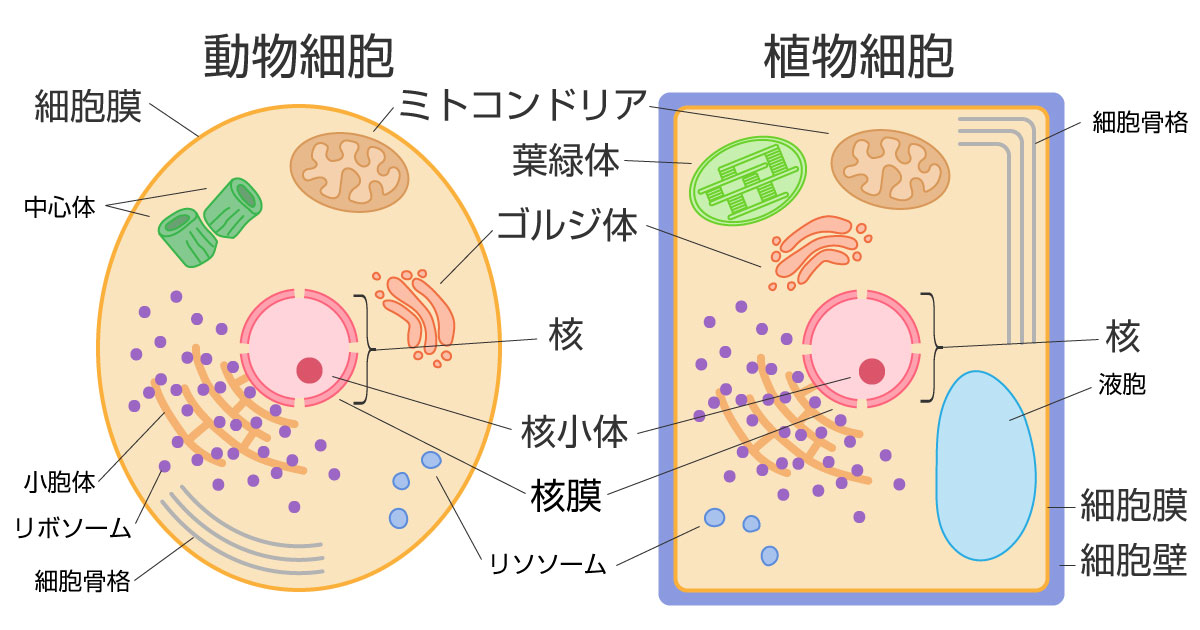
ちょっとごちゃごちゃしていますが、A、T、G、Cが作る暗号のような列(すなわちDNA配列)は、動物の場合には、核とミトコンドリアにあります。植物の場合には、核とミトコンドリアと葉緑体にあります(葉緑体は植物にしかありません)。以下では、人間の場合に限って話を進めます。
※DNA配列全体、あるいは核にあるDNA配列全体、ミトコンドリアにあるDNA配列全体、葉緑体にあるDNA配列全体を指したい時に、「ゲノム」という言葉を使います。ゲノムというのは、DNA配列全体という意味です。
人間の場合には、核とミトコンドリアにDNA配列がありますが、核にあるDNA配列とミトコンドリアにあるDNA配列は、長さが全然違います。核にあるDNA配列は、A、T、G、Cが3000000000文字(30億文字)並んでいるような感じです。ミトコンドリアにあるDNA配列は、A、T、G、Cが16000文字並んでいるような感じです。桁違いです。遺伝情報のほとんどは核にあると考えて間違いありません。
核の中にあるDNA配列は、完全に一続きになって存在しているわけではなく、いくつかに分かれて存在しています。人間の場合には、染色体と呼ばれる棒状の構造物が46本あり、それぞれにDNA配列の一部が存在するという具合です。46本の染色体は普段は絡まり合っており、細胞分裂が始まる時にはっきり分かれて見えます。人間の女性と男性は、以下の46本の染色体を持っています(図はDHC様のウェブサイトより引用)。
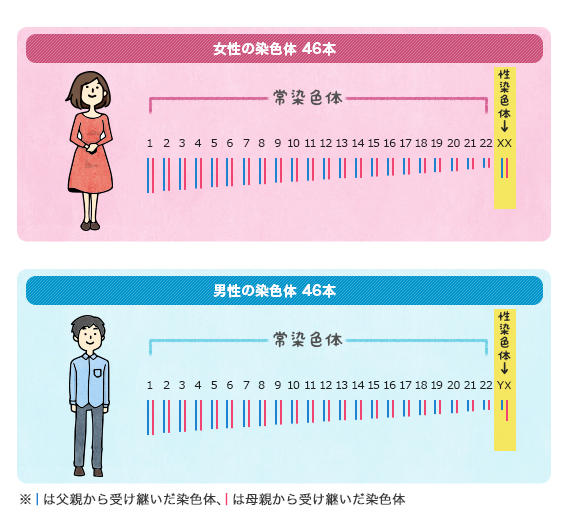
女性が持っているのは、

男性が持っているのは、

第1染色体~第22染色体のところに関しては、女性と男性で事情は同じです。残りの二本の染色体が「父からもらったX染色体と母からもらったX染色体」であれば、その人は女性になり、残りの二本の染色体が「父からもらったY染色体と母からもらったX染色体」であれば、その人は男性になるという仕組みです。XとXなら女の子になり、YとXなら男の子になるということです。
※第1染色体~第22染色体は、女性と男性に共通しているので、「常染色体」と呼ばれます。X染色体とY染色体は、性の決定に関わっているので、「性染色体」と呼ばれます。
すでに述べたように、遺伝情報のほとんどは核にあります。ミトコンドリアDNAは、核DNAに比べれば、微々たる存在です。Y染色体DNAは、核DNAの一部ですが、核DNAに占める割合は、上の図からもわかるように、小さいです。ミトコンドリアDNAとY染色体DNAを調べても、人間のDNA全体のごく一部を調べているにすぎないのです。
ミトコンドリアDNAが盛んに調べられてきたのは、ミトコンドリアDNAが母から娘へ(突然変異を除いて)代々不変的に伝わるからです。Y染色体DNAが盛んに調べられてきたのは、Y染色体DNAが父から息子へ(突然変異を除いて)代々不変的に伝わるからです。このような単純な特徴があるために、ミトコンドリアDNAとY染色体DNAは分析・解釈が比較的容易なのです。しかし、ミトンコンドリアDNAとY染色体DNAは例外的であり、DNAのほとんどはそのような単純な仕方で子孫に伝わりません。
●精細胞と卵細胞ができる減数分裂
人間の体で起きる細胞分裂には、体細胞分裂と減数分裂があります。このうちの減数分裂は特殊な分裂であり、しっかり理解しておく必要があります。
「減数分裂」という言い方は変じゃないかと思われる方がいるかもしれません。細胞が分裂したら細胞の数は増えるに決まっているからです。「減数分裂」の「減数」は、細胞の数が減るという意味ではなく、細胞の中に入っている染色体の数が減るという意味です。46本の染色体を持つ細胞から、23本の染色体しか持たない細胞ができるのです。この23本の染色体しか持たない特殊な細胞が、精細胞と卵細胞です。
精細胞のほうがわかりやすいので、精細胞のほうから説明します。46本の染色体を持つ精母細胞から、23本の染色体しか持たない精細胞ができます。もととなる精母細胞には46本の染色体が含まれていますが、いっぺんに46本に注目すると頭が混乱してしまうので、46本のうちの2本、ここでは父からもらった第1染色体と母からもらった第1染色体に注目しましょう。
(1)もととなる精母細胞には、父からもらった第1染色体と母からもらった第1染色体が入っています(本当は46本の染色体が入っていますが、今は2本だけを見ています)。
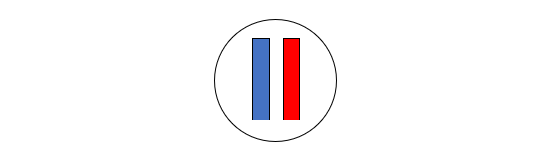
(2)父からもらった第1染色体は一本の棒状構造物でしたが、これが二本の棒状構造物になります。父からもらった第1染色体にあったDNA配列は複製されます。同様に、母からもらった第1染色体は一本の棒状構造物でしたが、これが二本の棒状構造物になります。母からもらった第1染色体にあったDNA配列は複製されます。
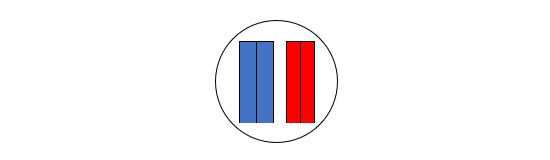
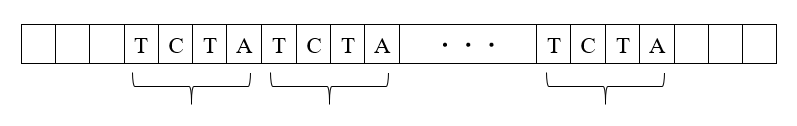
(3)左の二本の棒状構造物と右の二本の棒状構造物の間で、以下のようなDNA配列の部分的交換が行われます(ここに示したのは一つのパターンです)。
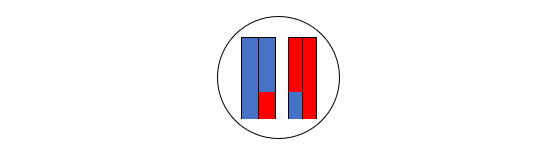
(4)左の二本の棒状構造物を含む細胞と、右の二本の棒状構造物を含む細胞に分裂します。
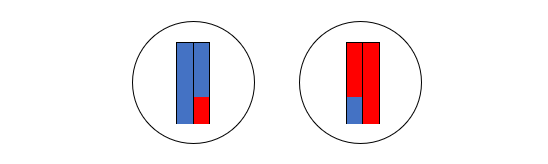
(5)これらの二個の細胞が、さらに分裂します。
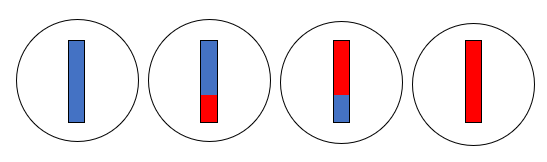
こうしてできた四個の細胞が、精細胞です。もととなった精母細胞には第1染色体が二本含まれていましたが、できた精細胞には第1染色体が一本しか含まれていません。
一応言葉で説明しましたが、(1)~(5)の図だけをさっと見たほうがわかりやすいかもしれません。DNA配列の複製、DNA配列の部分的交換、一回目の分裂、二回目の分裂という展開です。
ここでは第1染色体に注目しましたが、第1染色体に起きたのと同じことが第2染色体~第22染色体にも起きます。精細胞には、第1染色体が一本だけ、第2染色体が一本だけ、・・・、第22染色体が一本だけ含まれることになります。
もととなる精母細胞には一本のY染色体と一本のX染色体が含まれていますが、これらの間にはDNA配列の部分的交換はほとんど起きません。Y染色体がその端のわずかな領域を除いてDNA配列の部分的交換を行う能力を持っていないからです。したがって、精母細胞に含まれていたY染色体のDNA配列、または精母細胞に含まれていたX染色体のDNA配列がほとんどそのまま精細胞に含まれることになります。Y染色体とX染色体のペアは、大体(1)~(5)の図のようになるが、途中でDNA配列の部分的交換がほとんど起きないと理解しましょう。
第1染色体が一本、第2染色体が一本、・・・、第22染色体が一本、そしてY染色体またはX染色体が一本、計23本の染色体が精細胞に含まれることになります。
減数分裂は複雑すぎると感じる方は、(1)の図を見てこれが精母細胞、(5)の図を見てこれが精細胞、という理解でも十分です。(1)の段階では存在しなかった染色体が、(5)の段階では存在しているのがわかるでしょう。(5)の図の真ん中の二本がそうです。(5)の図の一番左の染色体が子に与えられるかもしれません。二番目の染色体が子に与えられるかもしれません。三番目の染色体が子に与えられるかもしれません。四番目の染色体が子に与えられるかもしれません。確率の問題です。
ポイントは、精細胞が作られる過程で、今まで存在しなかったDNA配列を持つ染色体が生じるということです。
精細胞が減数分裂を通じて作られるように、卵細胞も減数分裂を通じて作られます。ただし、一つの精母細胞から四つの精細胞ができますが、一つの卵母細胞から四つの卵細胞はできません。一つの細胞だけが卵細胞になり、残りの細胞は退化してしまいます。精細胞の場合と同様に、卵細胞には、第1染色体が一本だけ、第2染色体が一本だけ、・・・、第22染色体が一本だけ含まれることになります。
もととなる卵母細胞には二本のX染色体が含まれていますが、これらの間にはDNA配列の部分的交換が起きます。つまり、二本のX染色体には、二本の第1染色体、二本の第2染色体、・・・、二本の第22染色体と同じことが起きます。X染色体とX染色体のペアは、(1)~(5)の図のようになると理解しましょう。
第1染色体が一本、第2染色体が一本、・・・、第22染色体が一本、そしてX染色体が一本、計23本の染色体が卵細胞に含まれることになります。
ここでもポイントは、卵細胞が作られる過程で、今まで存在しなかったDNA配列を持つ染色体が生じるということです。
※減数分裂は、一見面倒くさいことをしているように見えるかもしれません。しかし、(5)の図で、旧来のDNA配列(外側の二つ)と新しいDNA配列(内側の二つ)が存在していることに注目してください。旧来のDNA配列を残しつつ、新しいDNA配列を生み出しているところに、巧妙さが感じられます。逆に、(1)の図で、旧来のDNA配列を残しつつ、新しいDNA配列を生み出すにはどうしたらよいか考えてみてください。そう考えると、(1)~(5)の手順が、非常に合理的で無駄がなく、最小手順に思えてきます。
●確かにDNA配列の部分的交換は起きるが・・・
上に説明したように、第1染色体~第22染色体にあるDNA配列およびX染色体にあるDNA配列は、そのまま不変的に子孫に伝わっていくものではありません。上の説明の(3)の段階で起きるDNA配列の部分的交換が重要です。(3)の図では、DNA配列の交換が一箇所で起きていますが、DNA配列の交換が複数の箇所で起きることもあります。しかしそれでも、せいぜい数箇所です。
核に含まれている各染色体は長いです。最も短い染色体でA、T、G、Cが5000万文字、最も長い染色体でA、T、G、Cが2億5000万文字並んでいるような感じです。このような染色体に存在する特定のとても短いDNA配列を考えてみましょう。
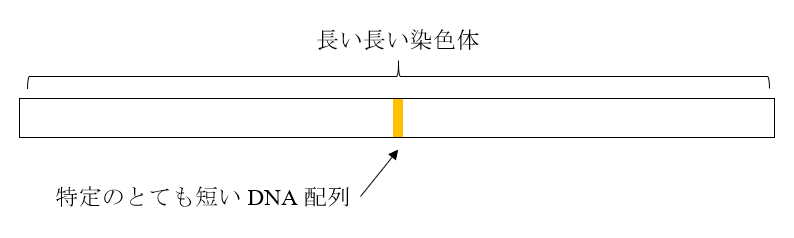
この染色体は、そのまま不変的に子孫に伝わっていくものではありません。(3)の図のような切断・再結合が行われ、そのような切断・再結合を経た染色体が子孫に伝わっていくこともあります。(5)の図を見ればわかるように、今まであったDNA配列を持つ染色体が子に与えられることもあれば、今までなかったDNA配列を持つ染色体が子に与えられることもあります。
察しがつくと思いますが、上の図の染色体の長さに対して特定のDNA配列の長さがとても短ければ(実際には、染色体自体がとてつもなく長いので、ある程度長くても大丈夫です)、その特定のDNA配列はなかなか切断されず、世代から世代へしぶとく残っていきます。染色体は(3)の図のように切断されることはありますが、千切りにされるわけではないからです。
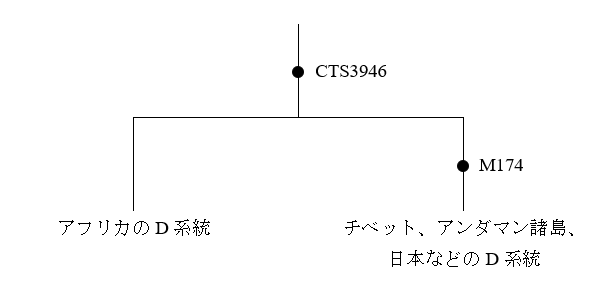
ミトコンドリアDNAとY染色体DNAは重要ですが、第1染色体~第22染色体とX染色体も捨てたものではないのです。ミトコンドリアDNAは、女→女→女→女→女という形でしか伝わらないという強い制限があります。Y染色体DNAは、男→男→男→男→男という形でしか伝わらないという強い制限があります。ミトコンドリアDNAとY染色体DNAだけを調べて、人類の歴史全体を窺おうとするのは無理があります。
実際、ミトコンドリアDNAとY染色体DNAを調べてもわからなかったが、DNA全体を調べてわかってきたこともたくさんあります。非常に重要な研究が蓄積してきたので、いくつか紹介することにしましょう。