モンゴル帝国の初代皇帝であるチンギス・ハーンは世界的に有名な人物ですが、彼とその男の子孫たちは大勢の子どもを残しました(Zerjal 2003)。北ユーラシアのY染色体DNAの分布にいくらか変化が生じるほどのインパクトがありました。
チンギス・ハーンたちの行動が異常だったのかというと、そういうわけではないようです。黄河文明が栄えていた頃に生きていたほんの三人の男性のY染色体DNAが、現代の中国人男性の40パーセントに受け継がれているという研究もあります(Yan 2014)。これもおそらく権力者からの拡散があったのでしょう。
筆者が古代から現代までのミトコンドリアDNAとY染色体DNAに関する膨大な研究を見て思ったのは、人類ではもともと、子どもづくりに参加する女性より子どもづくりに参加する男性が少なく、男性のほうに力関係(権力、武力)あるいは物質的豊かさの点で大きな差が存在すると、少数の男と多数の女による子どもづくりの傾向が顕著に強まるのではないかということでした。
上のチンギス・ハーンや黄河文明の権力者の例は、もともと人類にあった傾向が端的に現れた例ではないかというわけです。
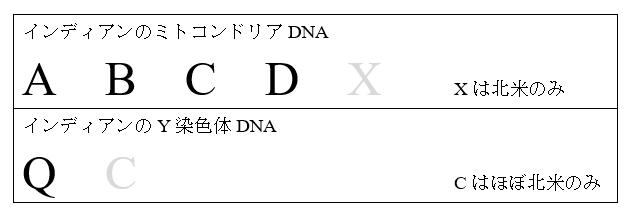
アメリカ大陸のインディアンのミトコンドリアDNAとY染色体DNAの話を思い出してください。アメリカ大陸のインディアンの研究には、古代北ユーラシアの姿を知るという重大な意味があります。アメリカ大陸のインディアンには、A、B、C、D、Xという五系統のミトコンドリアDNAとQ、Cという二系統のY染色体DNAが見られます(O’Rourke 2010)。この中で、ミトコンドリアDNAのX系統とY染色体DNAのC系統は、アメリカ大陸のどこにでも見られるわけではなく、分布が限られています。ミトコンドリアDNAのX系統は、北米の一部に低率で見られるだけです(Derenko 2001)。Y染色体DNAのC系統は、南米にもほんの少し存在しますが、やはりほぼ北米(特にアラスカとその近く)に限られています(Zegura 2004)。
南米のインディアンに限って見れば、ミトコンドリアDNAはA、B、C、Dの四系統、Y染色体DNAはほぼQの一系統のみという構成になっています。南米のインディアンは、古い時代にユーラシア大陸の北東部からアメリカ大陸に入っていった人々であると考えられるので(閉ざされていたアメリカ大陸への道の記事でお話ししたように、南米では14500年前頃から遺跡が見られ始めます)、南米のインディアンのミトコンドリアDNAとY染色体DNAは、古い時代のユーラシア大陸の北東部の状況をよく映し出していると見られます。
ミトコンドリアDNAのA系統、B系統、C系統、D系統とY染色体DNAのQ系統が支配し、その他の系統はあっても微々たる程度にすぎないという状況が、古い時代のユーラシア大陸の北東部に存在したと考えられます。ミトコンドリアDNAのA系統、B系統、C系統、D系統はどこからやって来たのでしょうか。Y染色体DNAのQ系統はどこからやって来たのでしょうか。
Y染色体DNAのQ系統については、一貫性を示す古代北ユーラシアの人々のDNAの記事で示したように、中東→中央アジア→バイカル湖周辺(さらに北ユーラシア全体)という流れに属することが明瞭なので、ミトコンドリアDNAのA系統、B系統、C系統、D系統がどこからやって来たのか考えましょう。
●ミトコンドリアDNAのM系統とN系統から考える
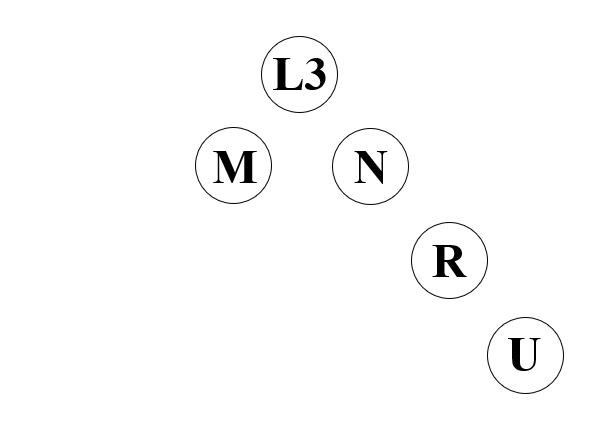
アフリカに見られるミトコンドリアDNAのL3系統から、アフリカの外に見られるM系統とN系統が生まれたことは、すでに述べました。M系統とN系統というのは巨大なくくりで、アフリカの外に見られるミトコンドリアDNAはすべてM系統かN系統のどちらかに属します。A系統とB系統はN系統のほうに属する下位系統で、C系統とD系統はM系統のほうに属する下位系統です。
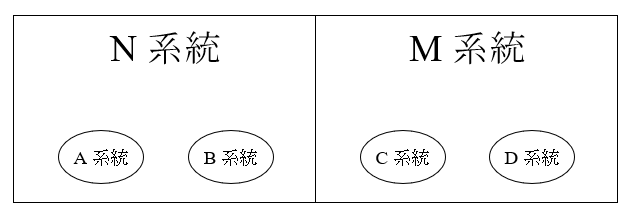
ミトコンドリアDNAのN系統とM系統がアフリカの外の世界にどのように分布しているか見ると、大変興味深いことがわかります。N系統はアフリカの外の世界のどこにでもよく見られますが、M系統はアフリカの外の世界で非常に偏った分布を見せています。M系統は南アジアおよびそれより東(つまり東アジア・東南アジアを含む東ユーラシア、南北アメリカ大陸、オセアニア)に偏在しているのです。
例外的なのは特にアフリカの東部に見られるM系統(より詳しくはM1系統)ですが、アフリカのM系統はアジアのM系統ほどの深い歴史は持っておらず、アジアに存在したM系統の一部がアフリカに戻ったと考えられています(González 2007)。Initial Upper Paleolithic(後期旧石器時代の初期)の話の中でU系統の一部(U6系統)が中東からアフリカに戻ったことをお話ししましたが(Olivieri 2006)、同じようにM系統の一部(M1系統)も中東からアフリカに戻ったということです。
※人類が進出してまもない頃のヨーロッパでミトコンドリアDNAのM系統が少し見られたこともありましたが、その後消滅してしまいました(Posth 2016)。
アフリカの外の世界のどこにでもよく見られるN系統と違い、連続的でありながら限定的な分布の仕方をしているM系統は、どのように拡散したかが見通しやすいです。M系統は以下のように拡散したと考えられます。
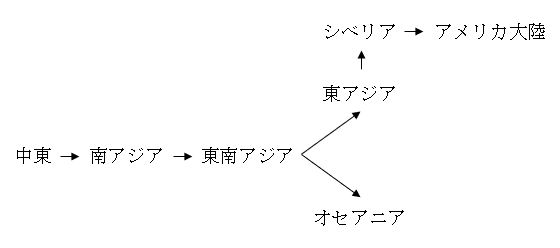
ミトコンドリアDNAのC系統とD系統がM系統に属する下位系統であることはすでに述べました。このことからつまり、古い時代にユーラシア大陸の北東部に存在したC系統とD系統は、中東→南アジア→東南アジア→東アジア→シベリアという流れに属するものであるということになります。古い時代のユーラシア大陸の北東部で支配的だったA系統、B系統、C系統、D系統のうちの少なくともC系統とD系統は、中東→南アジア→東南アジア→東アジア→シベリアという流れに属するということです。
A系統とB系統はどうでしょうか。N系統に属するA系統とB系統に関しては、C系統とD系統の場合のように容易に判断することはできません。一貫性を示す古代北ユーラシアの人々のDNAの記事で示したように、N系統は中東から北ユーラシア方面にも、パプアニューギニア・オーストラリア方面にも拡散しています。古い時代のユーラシア大陸の北東部に存在したA系統とB系統は、北側ルート(中央アジア経由)の流れに属するものかもしれないし、南側ルート(東南アジア経由)の流れに属するものかもしれません。
一貫性を示す古代北ユーラシアの人々のDNAの記事でお話ししたように、日本の近辺で発見された現生人類として最も古い北京郊外の田園洞遺跡で発見された4万年前の男性は、B系統のミトコンドリアDNAを持っていました。4万年前の東アジアの記事でバイカル湖方面からやって来る人々と中国南部・東南アジア方面からやって来る人々が混ざり合う話をしましたが、田園洞の男性がいたのはちょうどそのあたりで、田園洞の男性のB系統のミトコンドリアDNAが北側ルート(中央アジア経由)の流れに属するのか、南側ルート(東南アジア経由)の流れに属するのかきわどいところです。長くなるので、ここでいったん切ります。
参考文献
Derenko M. V. et al. 2001. The presence of mitochondrial haplogroup X in Altaians from South Siberia. American Journal of Human Genetics 69(1): 237-241.
González A. M. et al. 2007. Mitochondrial lineage M1 traces an early human backflow to Africa. BMC Genomics 8: 223.
Olivieri A. et al. 2006. The mtDNA legacy of the Levantine early Upper Palaeolithic in Africa. Science 314(5806): 1767-1770.
O’Rourke D. H. et al. 2010. The human genetic history of the Americas: The final frontier. Current Biology 20(4): R202-R207.
Posth C. et al. 2016. Pleistocene mitochondrial genomes suggest a single major dispersal of non-Africans and a Late Glacial population turnover in Europe. Current Biology 26(6): 827-833.
Yan S. et al. 2014. Y chromosomes of 40% Chinese descend from three Neolithic super-grandfathers. PLoS One 9(8): e105691.
Zegura S. L. et al. 2004. High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of Native American Y chromosomes into the Americas. Molecular Biology and Evolution 21(1): 164-175.
Zerjal T. et al. 2003. The genetic legacy of the Mongols. American Journal of Human Genetics 72(3): 717-721.