

●ミトコンドリアDNAのB系統
ミトコンドリアDNAのB系統の研究は、異なる表記と意見が飛び交って、大変わかりづらくなっています。現在では、ミトコンドリアDNAのB系統をB4系統、B5系統、B6系統の三つに分けて記述するのが標準的です。以下のピンク色の部分は内容が非常に専門的なので、最近の研究事情を詳しく知りたい方以外は読み飛ばして次に進んでください。
B1~B3という表記はもう基本的に使われず、アメリカ大陸のインディアンのミトコンドリアDNAを記述する時に昔のB2という表記を用いることがあるくらいです。昔のB2は今ではB4系統の下位系統として分類されています。B6系統(150、8281-8289d、9452、12950、13928Cという変異によって定義される)は、研究者によってB6と表記したり、B7と表記したりしています。
表記だけでなく、意見もばらついています。以前にお話ししたようにミトコンドリアDNAのN系統の一下位系統としてR系統があり、R系統の一下位系統としてB系統があります。現在特に問題なのは、(1)のような見方をする研究者(Kong 2003など)と、(2)のような見方をする研究者(Soares 2009など)がいることです。

(1)はB4とB5とB6が近い系統関係にあり、この三者とR11が遠い系統関係にあるという見方です。(2)はB4とB5が近い系統関係にあり、B6とR11が近い系統関係にあり、前の二者と後の二者が遠い系統関係にあるという見方です。(1)が従来の見方で、(2)はそれを変形する見方です。B4とB5とB6に8281-8289dという変異が共通し、B6とR11に12950という変異が共通しているために、このようなことになっています。大きく見れば、B4、B5、B6、R11はどれもR系統の下位系統であり、B4、B5、B6、R11に系統関係があることは間違いありません。近い系統関係がどこにあるかという点で、見方が分かれています。以下では、基本的に(1)にしたがって話を進めます。ただし、(1)であっても(2)であっても、ここでの話の大筋にはさほど影響ありません。
B系統のミトコンドリアDNAは東アジア・東南アジアを中心に非常に多く見られますが、そのほとんどはB4系統かB5系統です。ちなみに、B4系統の一下位系統がアメリカ大陸に入っていきました。そのB4系統とB5系統の陰に隠れて、B6系統は全くと言ってよいほど注目されてきませんでした。B6系統は、日本列島と朝鮮半島はもちろんのこと、中国でもなかなか見られませんが、東南アジア、特に南アジアから東南アジアへの入口の近く(つまりミャンマーのあたり)に向かっていくとだんだんと見られるようになります(Summerer 2014)。Summerer氏らの調査では、327名のミャンマー人のミトコンドリアDNAを調べて、21名(6.4%)がB6系統のミトコンドリアDNAを持っていました。これは、まずまず見られると言ってよいレベルでしょう。B系統の中で、B6系統は超少数派ですが、独自の変異をたくさん積み重ねており、とても古い時代(B系統の発生から長くは経っていない頃)にB4系統とB5系統と分かれていることは確実です(Kong 2003、Summerer 2014)。B4系統とB5系統の広大な分布とは違うB6系統の独特な分布は、大変示唆的です。
ミトコンドリアDNAのB系統が北側ルート(中央アジア経由)で東アジアに入ってきたと仮定すると、かなり不自然なことになります。B6系統は北方には見られず、南下してもなかなか見られず、南アジアから東南アジアへの入口の近くに向かっていくとだんだんと見られるようになります。しかも、そのB6系統の歴史はとても古いのです。ミトコンドリアDNAのB系統はとても古い時代に南方で生じ、B系統の一部は北上し、一部は北上しなかったと考えるほうがよく合います。
今のところ、ミトコンドリアDNAのB系統は南側ルート(東南アジア経由)の流れに属する可能性が高そうです。B系統はR系統の一下位系統なので、R系統の他の下位系統、特にB系統に近そうな下位系統と照らし合わせながら根拠を固める必要があるでしょう。
●ミトコンドリアDNAのA系統
B系統以上に謎めいているのがA系統です。ミトコンドリアDNAのN系統にある変異が起きてR系統が生まれ、N系統にそれとは違う変異が起きてA系統が生まれました。下の図のように、B系統はR系統に属しますが、A系統はR系統に属しません。
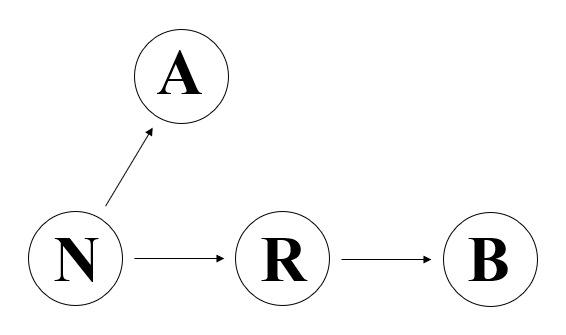
謎めいているのは、N系統からA系統が生まれる過程です。N系統と比べると、A系統は152、235、523-524d、663、1736、4248、4824、8794、16290、16319のようにとても多くの変異を起こしており、N系統からA系統に至るまでの道が非常に長いことがわかります(Kong 2003)。上に並べた一連の変異は、なんらかの順序で起きていったものです。上に並べた変異のうちの一つだけが起きたタイプ、上に並べた変異のうちの二つだけが起きたタイプ、上に並べた変異のうちの三つだけが起きたタイプ、・・・・・、上に並べた変異のうちの九つだけが起きたタイプのミトコンドリアDNAだってあったはずです。しかし、そのようなミトコンドリアDNAは見当たりません。つまり、A系統に近い系統はことごとく消滅し、かろうじてA系統だけが残ったのです。
N系統からA系統に至るまでの過程で順々に生まれたはずの系統がことごとく消滅したことは疑いなく、問題はそのような消滅を引き起こす(厳しい)状況が北のほうにあったのか、南のほうにあったのかということです。シベリアとシベリアを襲ったLast Glacial Maximum(最終氷期最盛期)が系統の大減少を引き起こした可能性は十分にあります。上に並べた10個の変異を持つA系統自体も、分布の中心が東ユーラシアの南のほうにはなく、東南アジアの一部にかすかに届いているだけなので、南のほうで生じたものではないと見られます(Stoneking 2010)。
今のところ、断定はできませんが、A系統は北側ルート(中央アジア経由)の流れに属する可能性が十分にあります。ことごとく消滅してしまった「ゴースト系統」(N系統からA系統に至るまでの過程で生じた系統)のミトコンドリアDNAを持つ古代人が北ユーラシアで発見されれば、その可能性が強まります。
前回の記事と今回の記事を整理すると、以下のようになります。
インディアンのミトコンドリアDNA
A系統 — 北側ルートの流れに属する可能性が十分にある
B系統 — 南側ルートの流れに属する可能性が高い
C系統 — 南側ルートの流れに属する
D系統 — 南側ルートの流れに属する
インディアンのY染色体DNA
Q系統 — 北側ルートの流れに属する
インディアンのミトコンドリアDNAを見ると、東南アジア方面からの流れが強そうなのに、インディアンのY染色体DNAを見ると、中央アジア方面からの流れが圧倒的なのです。
アフリカ・中東からインディアンの先祖がいたと考えられるユーラシア大陸北東部に辿り着くためには、東南アジアを経由するか、中央アジアを経由するかしなければなりません。インディアンの先祖は、東南アジア方面から来たのか、中央アジア方面から来たのか、あるいは両方面から来たのか、もしそうなら東南アジア方面から来た人が多かったのか、中央アジア方面から来た人が多かったのかと議論されてきたのは当然です。しかし、上のようなインディアンのミトコンドリアDNAとY染色体DNAの対照的な傾向を見ると、男女関係も考えないわけにはいきません。
ここで二つの問題が持ち上がります。一つ目の問題は、インディアンのミトコンドリアDNAとY染色体DNAはなぜ対照的な傾向を示しているのかという問題です。二つ目の問題は、インディアンのミトコンドリアDNAとY染色体DNAが示している対照的な傾向はインディアンに特有なものなのか、つまりインディアンに限定されたものなのかという問題です。
参考文献
Kong Q.-P. et al. 2003. Phylogeny of east Asian mitochondrial DNA lineages inferred from complete sequences. American Journal of Human Genetics 73(3): 671-676.
Soares P. et al. 2009. Correcting for purifying selection: An improved human mitochondrial molecular clock. American Journal of Human Genetics 84(6): 740-759.
Stoneking M. et al. 2010. The human genetic history of East Asia: Weaving a complex tapestry. Current Biology 20(4): R188-R193.
Summerer M. et al. 2014. Large-scale mitochondrial DNA analysis in Southeast Asia reveals evolutionary effects of cultural isolation in the multi-ethnic population of Myanmar. BMC Evolutionary Biology 14: 17.